 |
Новости |  |
|
Библиотека | |
|
Новые книги | |
|
Ссылки | |
|
Карта сайта | |
|
О нас | |
 |

Микроклимат гнездаТерморежимТерморецепторы. Температура среды в известной степени влияет на все биохимические и биофизические процессы, протекающие в организме. Поэтому на изменение температуры среды реагирует все тело пчелы. Влиянию температуры подвержена также работа различных узкоспециализированных рецепторов, даже таких, как фоно- и хеморецепторы. Специализированные температурные рецепторы животных разделяются на две категории — холодовые и тепловые. Как те, так и другие реагируют на колебания температуры изменением количества генерируемых импульсов. Благодаря этому они являются индикаторами изменения температуры. При этом холодовые рецепторы отвечают временным возрастанием импульсации на понижение температуры. С ее повышением импульсация этого типа рецептора уменьшается или даже полностью затормаживается. Тепловой рецептор, напротив, реагирует увеличением количества импульсов на нагрев. У пчел терморецепторы холода представляют собой ампулоподобные сенсиллы длиной 10—20 мкм и шириной до 10 мкм, расположенные на восьми дистальных члениках антенн. Наибольшей чувствительностью к температуре обладают, по данным поведенческих опытов X. Херана, терморецепторы, расположенные на первых пяти дистальных члениках. В электрофизиологических экспериментах В. Лачера (1964) показано, что резкая активизация рецептора, выражающаяся в увеличении количества генерируемых им импульсов, наблюдается лишь в течение непродолжительного времени непосредственно после его охлаждения. Так, на понижение температуры с 26° до 23° рецептор отвечает почти десятикратным увеличением частоты пульсаций. Но уже через 1 с количество генерируемых им импульсов уменьшается примерно вдвое. Результаты этих электрофизических исследований свидетельствуют о способности пчел воспринимать колебания температуры, составляющие десятые доли градуса. Действительно, понижение температуры на 0,5 °С отражается на увеличении импульсации холодового терморецептора рабочей пчелы на один импульс в секунду. В тех же пределах находится чувствительность терморецепторов трутней: при понижении температуры на 0,25° зарегистрировано увеличение частоты пульсаций на 0,5 Гц. Нагрев рассматриваемых рецепторов стимулирует понижение их активности. Полное затухание пульсаций рецептора наблюдается, когда температура среды достигает 40—45 °С. Это подтверждает то, что описываемый орган является типичным холодовым рецептором. Типичных рецепторов тепла у пчел не обнаружено. Их функцию совмещают рецепторы углекислого газа, реагирующие на нагрев увеличением частоты генерируемых импульсов. Эти рецепторы представляют морфологически ампуловидные или целоконические сенсимы, расположенные, как и холодовые рецепторы, на девяти дистальных члениках антенн. Механизмы теплогенерации и охлаждения тела пчел. У пчел, как и у других пойкилотермных животных, температура тела зависит в значительной мере от температуры окружающей среды. Но наличие этой зависимости не означает равенства или некоторого постоянного соотношения между этими температурами. В частности, по данным Е. Шульз-Лангер (1958), разность между температурой тела летающих пчел и среды уменьшается с девяти до одного градуса при увеличении температуры окружающего их воздуха с 9° до 34 °С. Это свидетельствует о том, что пчелы регулируют в некоторых пределах температуру своего тела. Механизм производства тепла у пчел основан на мышечной активности. Наибольшее его количество выделяется грудной мускулатурой. Об этом говорит тот факт, что разогрев пчелы начинается с подъема температуры груди. По окончании периода разогрева (как и в процессе его) температура груди выше, чем брюшка. Роль летательных мышц непрямого действия как генераторов тепла доказана в электрофизиологических экспериментах X. Эша (1964). Исследователь регистрировал одновременно температуру в груди пчелы, вибрации ее зкзоскелета (хитинового покрова) под действием мышц непрямого действия и электрические процессы в этих мышцах. В результате обнаружено, что выделение тепла всегда сопровождается генерацией потенциалов действия. С увеличением частоты их следования происходит подъем температуры груди. При небольшой частоте следования потенциалов (до 10 Гц) процесс тепловыделения не сопровождается вибрациями экзоскелета груди или следует в виде серии повторяющихся импульсов. Вибрации груди, приводящие крылья в колебательные движения (как это делают при полете пчелы), и значительное повышение температуры груди наблюдается в тех случаях, когда потенциалы действия следуют с частотой выше 10 Гц. В основном за счет работы грудной мускулатуры повышается температура тела пчелы, готовящейся к полету. Поэтому при подготовке к полету сильнее всего разогревается грудь насекомого. Значительно слабее — брюшко. Так, при температуре среды около 18 °С температура груди пчелы перед взлетом от улья поднимается до 33—36 °С. В то же время температура брюшка изменяется примерно на 2°С и достигает 21—22 °С. Скорость разогрева пчелы, готовящейся к полету, составляет, по данным X. Эша (1960), 2,1±0,25 °С в мин. Температура тела пчелы, закончившей полет, на 6— 20 °С выше температуры окружающей среды. При этом температура груди бывает на 5—11°С выше температуры брюшка. Разогрев тела пчелы при полете зависит от внешней температуры. В частности, температура тела пчелы при температуре 22—26 °С достигает 35—37 °С (X. Эш, 1960), а при 35—37 °С нагревается до 42 °С (Е. Шульз-Лангер, 1958). Интересное явление обнаружил X. Эш (1960), регистрировавший температуру тела пчел, прилетающих к кормушке с раствором сахара. Оказывается, у пчел, впервые прилетающих к кормушке, температура тела примерно на 1 °С ниже, чем у тех, которые длительное время посещают этот источник пищи. Указанное различие сглаживается после 4—5 визитов к кормушке. Значительно возрастает температура тела пчел при повышении их двигательной активности. Быстрый ее подъем происходит у пчел, проявляющих интерес к особям, которые информируют членов семьи об обнаруженном источнике пищи. Передача этой информации происходит в процессе передвижения сигнализирующих и мобилизуемых сигнальщицами пчел. У последних за 2,2 мин. температура тела может подняться на 6°С, достигнув 39—40 °С (X. Эш, 1960). Теплогенерация пчел не обязательно связана с повышением их двигательной активности. У внешне неподвижных пчел может происходить быстрый подъем температуры груди. В таком случае тепло образуется в результате микроколебаний грудных мышц непрямого действия (мышцы летательного аппарата), что подобно явлению дрожи у млекопитающих. Разогрев тела пчел возможен также за счет поглощения ее покровами тепловой энергии, в том числе солнечной. Это особенно проявляется в жаркую погоду. Например у пчел, летающих при температуре 32—34 °С под открытыми лучами солнца, температура тела бывает примерно на 4° выше, чем у насекомых, летающих при тех же условиях, нов тени (Е. Шульз-Лангер, 1958). В связи с тем, что тело пчелы обладает высокой теплопроводностью и соответственно низкими теплоизоляционными свойствами, нагретая пчела быстро охлаждается. Так, наркотизированная пчела, нагретая в инфракрасных лучах до 40 °С, охлаждается при температуре 22 °С до 30 °С примерно за 35 с. Дальнейшее охлаждение до 25 °С происходит за 60—70 с (X. Эш, 1960). Отсюда следует, что отдача тепла нагретой пчелой происходит по экспоненциальному закону с постоянной времени около 70 с. Некоторое количество тепла расходуется в процессе испарения. Основную часть составляет испарение метаболической воды, выделяемой через дыхальца. Пожалуй, больше всего происходит охлаждение пчелы за счет этого процесса во время полета, когда она расходует по отношению к другим формам деятельности значительное количество корма. Но и в этом случае охлаждение за счет испарения не превышает 10% от общей суммы теплопотерь (X. Эш, 1960). Что касается кутикулярного испарения, то оно, по всей вероятности, не может играть существенного значения в механизме охлаждения пчелы, поскольку критическая температура воска, при которой он имеет наибольшую водопроницаемость, находится значительно выше витальной для пчел. Важную роль в регуляции температуры тела пчел играет их способность изменять в широких пределах интенсивность обмена веществ. Действительно, интенсивность дыхания пчел, выражающаяся в потреблении ими кислорода, растет с повышением внешней температуры до 30 °С. При дальнейшем ее повышении потребление кислорода уменьшается. Это может служить объяснением того, что разогрев тела пчел прекращается или значительно ослабевает, когда температура среды достигает указанного значения. Сезонные и суточные колебания температуры в гнезде. Температура тела пчелы, находящейся вне гнезда, может колебаться в зависимости от ее состояния и условий среды в широких пределах. В отличие от этого температура в пчелином гнезде поддерживается членами семьи с довольно высокой стабильностью, особенно в зоне расплода. Ее верхняя граница при относительно высокой внешней температуре редко поднимается выше 36° С. Уместно подчеркнуть, что семья медоносных пчел, лучше других, близких к ним по образу жизни общественных насекомых, может противостоять действию высоких внешних температур. На это указывают, например, данные Л. Верма (1970), проводившего в одно и то же время измерения температур в гнездах медоносных и индийских пчел. Оказалось, что при температуре внешнего воздуха в дневные часы 4 Г С, максимальная внутригнездовая температура у первых составляла 36,5°, у вторых —38° С. На терморежим в пчелином жилище в зоне расположения расплода заметное влияние оказывает температура внешней среды. При повышении внешней температуры с 5Р до 27° С температура в зоне пчелиного расплода семей грузинской и среднерусской рас увеличивается в среднем с 33,5—34,5° до 36—36,3° С. Абсолютное значение и стабильность температуры зависят от места расположения расплода. В течение весенне-летнего периода развития семьи на одной и той же рамке с разновозрастным пчелиным расплодом наиболее высокая и стабильная температура бывает в центральной зоне гнезда. Здесь слабо или совсем не прослеживается влияние суточных колебаний внешней температуры. Менее чем на 1°С изменяется среднечасовая температура в этой зоне гнезда при суточных внешних колебаниях с 26° до 16° С. Среднее значение температуры в этой зоне находится на уровне 35° С. Такая температура поддерживается в зоне расплода, расположенного на расстоянии 5—7 см от летка в диагональном направлении к центру рамки. Данные непрерывной регистрации температуры в центральной зоне гнезда с помощью микротермодатчика (типа МТ-54), говорят о том, что на короткое время (несколько минут) температура воздуха в центральной части гнезда может опускаться до 29—30° С. Это наблюдается чаще всего в дневные часы, когда большое количество пчел вылетает из улья. Иногда такие понижения температуры воздуха регистрируются и в ночные часы. Важно отметить, что во время этих скачкообразных понижений температуры воздуха находящийся поблизости расплод не успевает охладиться. Относительно низкую температуру поддерживают пчелы на расплоде, расположенном на периферии гнезда. Так, на центральной рамке в ее верхнем углу у стенки, противоположной летковому отверстию, средняя температура составляет 33,5° С. Ее абсолютное значение прямо связано с внешней температурой. Суточные колебания температуры в пределах 10° С отражаются на ее изменении в указанной зоне гнезда семьи, состоящей из 26—29 тыс. взрослых особей и 12—13 тыс. ячеек с расплодом, более чем на 1,5° С. Особенно значительное понижение температуры бывает в этой зоне гнезда при длительных летних похолоданиях, когда в течение нескольких часов (иногда 2—3) она может удерживаться на уровне 28,5—29° С. Понижение температуры в зоне расплода можно вызвать, отделив в течение нескольких часов большое количество пчел. Так, при потере семьей примерно 15 тыс. рабочих особей температура в зоне расплода снижается на 2—3°С. Анализ показаний температуры за весенне-летний сезон в зоне пчелиного расплода семей среднерусской и серой горной кавказской расы свидетельствует о том, что она отличается в них в основном при относительно низкой внешней температуре. Так, когда температура внешней среды находится на уровне 5—14° С, в расплодной части гнезда семьи среднерусской расы она составляет в среднем 34,4—35° С (см. рис. За), а в семье серой горной кавказской расы — 33,7—34,7° С. Высокой стабильностью характеризуется температурный режим в зоне выращивания маток. У роевых и свищевых маточников температура слабо подвержена колебаниям внешней температуры. Например, на понижение внешней температуры с 23° до 11° С температура вблизи маточника опускается в среднем не более чем на 0,5° С. Подобно тому как это имеет место в зоне пчелиного расплода, температура у маточника независимо от внешней может кратковременно опускаться до 28—29° С. Эти понижения температуры не имеют выраженной цикличности и не связаны с какой-либо определенной фазой развития маток. Средняя температура у маточника 34° С. Что касается трутневого расплода, то пчелы слабо заботятся о его терморежиме. Так, при внешней температуре до 20° С температура в зоне трутневого расплода бывает на 1—2° С ниже, чем в области выращивания рабочих пчел. Это различие уменьшается до 1—0,5° С, когда температура внешнего воздуха достигает 25—26° С. К концу лета количество расплода, выращиваемого семьей, постоянно уменьшается. Это связано с погодными условиями и наличием цветущих медоносных растений. В тропиках и субтропиках на протяжении всего года идет обновление состава семьи, а в полосе с умеренным климатом к началу — середине осени пчелы прекращают выращивать расплод. Его появление в этот период может быть вызвано значительным потеплением, особенно если этому сопутствует цветение медоносных растений, или искусственной стимуляцией активности семьи. По окончании развития семьи (после прекращения выращивания расплода) температура в гнезде, в том числе и в его центральной части, понижается. Однако если в это время в семье появится расплод (естественным путем или его перенести из другой семьи, не закончившей развитие), то терморежим в гнезде вновь стабилизируется. Это происходит за счет повышения активности рабочих пчел. С уменьшением расплода в гнезде сокращается зона с относительно стабильной температурой. Ее колебания за пределами области выращивания расплода бывают наибольшими в то время, когда днем настолько тепло, что пчелы еще занимаются внеульевыми работами, например, приносят цветочную пыльцу, прополис и т. п., а значительные ночные понижения температуры побуждают их собираться в небольшой зоне жилища. В связи с этим в течение суток сильнее всего изменяется температура в тех местах жилища, которые пчелы покидают в ночные часы. Днем температура здесь может достигать 30—35° С, а ночью опускаться в некоторых зонах улья, например в его нижней части, противоположной летковому отверстию, и у боковых стенок — до внешней. К рассматриваемому периоду относится формирование пчелами так называемого зимнего клуба. Его образуют пчелы, собравшиеся в относительно небольшой зоне жилища. Конструктивной особенностью зимнего клуба, отличающей его от роевой грозди, является то, что он разделен рядами сот. Лишь небольшое число пчел выходит в каком-либо месте, за пределы сот, образуя связующую часть клуба, которая перемещается обычно за период зимовки от передней стенки улья к надрамочному пространству. При очень сильных морозах эти пчелы могут временно уходить в глубь межрамочных пространств. В таком случае прямое контактирование между пчелами, разделенными рядами сот, прекращается. Большая часть пчел зимнего клуба формируется на пустых ячейках сот. Заполненные медом ячейки занимает только небольшая часть клуба. Потребление меда стимулирует пчел перемещаться вверх. Это связано также со стремлением разместиться в наиболее теплой части жилища. Действительно, в сильные морозы большая часть пчел, зимующих в ульях под открытым небом, располагается в верхних частях сотовых рамок. Но в ульях, донные части которых оборудованы электрическими нагревателями, поддерживающими температуру на уровне 7—10° С, клуб смещен к нижней части сотовых рамок. В таких ульях перемещение клуба вверх происходит при потеплениях и к весне. Стремлением пчел занять наиболее теплую зону в жилище можно объяснить также то, что при наличии в одном улье двух семей, разделенных сплошной перегородкой, клуб каждой из них формируется осенью у этой перегородки, как наиболее темной стенке жилища. Наиболее высокая и стабильная температура поддерживается в центральной или близкой к геометрическому центру зоне зимнего клуба, называемой тепловым центром. Она зависит от количества пчел в клубе, их физиологического состояния и внешней температуры. Обычно чем активнее пчелы, тем больше зона с высокой температурой. При похолоданиях температура в центральной зоне клуба возрастает. Это происходит непосредственно после понижения внешней температуры. С удалением от теплового центра зимнего клуба к его периферии температура постепенно снижается. В различных зонах периферической части клуба она может довольно долго поддерживаться на одном и том же уровне. Это бывает большей частью в тех случаях, когда пчелиные семьи зимуют в помещениях при постоянной температуре. Всякому перемещению клуба предшествует повышение пчелами температуры. В таких случаях может происходить перемещение теплового центра не только в вертикальной, но и горизонтальной плоскости. Последнее объясняется перераспределением количества пчел в межсотовых пространствах. К середине—концу зимы активность пчел повышается. Изменяется температура в клубе, расширяется область, занимаемая тепловым центром, стабилизируется температура в ней. С этого времени в семье обычно появляется расплод. Его количество с приближением весны увеличивается. Дальнейшее изменение температурного режима в пчелином жилище представляет по существу процесс, обратный тому, который наблюдается в конце лета — начале осени. Механизм терморегуляции, используемый семьей. Широкий ареал распространения медоносных пчел связан с тем, что в процессе эволюции общественного образа жизни они приспособились коллективными усилиями регулировать микроклимат своего гнезда. Благодаря этому пчелиная семья в состоянии жить в условиях, где диапазон годовых колебаний температур достигает почти 100°. Действительно, пчелиная семья выдерживает плюс 40—45° С и выживает в тех случаях, когда температура в период зимовки опускается до минус 50° С. Механизм терморегуляции пчелиной семьей представляет цепь сложных поведенческих актов, выполняемых рабочими особями. При этом они пользуются различны и средствами в зависимости от повышения или понижения по отношению к оптимальной температуре среды. Реакция на перегрев. Отрицательное отношение пчел к перегреву их жилища проявляется в естественных условиях еще при выборе мест для жилья. Так, если рою предоставить возможность такого выбора, то он поселится при прочих равных условиях в сооружении, защищенном от солнца. Следовательно, пчелы заботятся об экономии затрат энергии на регулирование микроклимата жилища. Выбор места для жилья не гарантирует семью от возможного перегрева. Поэтому в процессе эволюции пчелы приспособились активно противодействовать перегреву гнезда. Прежде всего перегрев побуждает пчел аэрировать жилище, для чего они машут крыльями. Частота взмахов и сопутствующих им звуков зависит от того, насколько температура превосходит оптимальную. Это прослежено на семьях, живших в ульях, донная часть которых была покрыта пластиной, нагреваемой электрическим током. (Е. Еськов, 1979). Оказалось, что при повышении температуры в нижней части гнезда с 33,1—35,5° до 37,0—42,0° С, частота звуков, издаваемых пчелами-вентилировщицами, возросла с 75—85 до 145—155 Гц (рис. 1). Усилением активности аэрации пчелами гнезда можно объяснить то, что несмотря на значительный перегрев периферической части гнезда, ее рост в центральной зоне — там, где находится расплод, составил всего 1,8° С. При этом температура на периферической части гнезда возросла на 4,7—9,1° С. 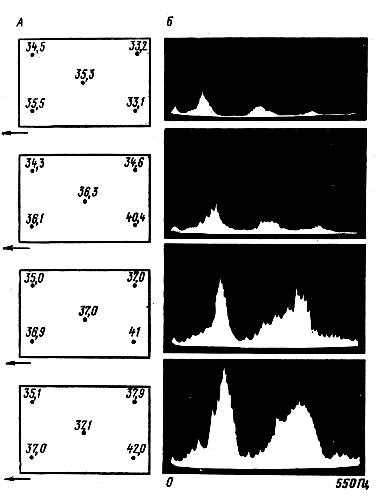 Рис. 1. Влияние разогрева гнезда на интенсивность и спектральную структуру звуков пчелиной семьи: А - температура, регистрируемая пятью термодатчиками, расположенными на средней рамке улья (стрелкой показано направление к летковому отверстию); Б — спектры звуков зарегистрированных в соответствующие моменты разогрева гнезда (по Е. Еськову, 1979) Звук пчел-вентилировщиц, являясь сопутствующим процессом, выполняет тем не менее важную сигнальную функцию, Оказывается, звуки, издаваемые вентилирующими особями, воспринимаются членами их семей как сигналы неблагополучия. Они побуждают других пчел принимать меры по защите гнезда от перегрева. Действительно, звуков, издаваемых особями, аэрирующими жилище, достаточно, чтобы при относительно высокой температурь стимулировать аналогичную реакцию у членов семьи, которые находятся в это время в любой части жилища. Пчел можно побудить к вентилированию не только в ответ на воспроизведение естественных звуков, сопутствующих процессу аэрации жилища, но и на чистые тона определенных частот. Для этого нужно поднести к голове спокойно сидящей пчелы отверстие трубки, соединенной со звуковым излучателем. При этом с повышением частоты (в диапазоне звуков, издаваемых самими пчелами-вентилировщицами) звука эффективность его действия на пчел усиливается. Наибольшей эффективностью действия обладают звуки частотой 140—160 Гц. Звуки такой частоты издают пчелы-вентилировщицы при значительном перегреве жилищ. Следовательно, чем больше опасность перегрева гнезда, тем активнее пчелы занимаются его аэрацией. В свою очередь, рост частоты звука вентилировщиц вовлекает в процесс аэрации жилищ все новых членов семьи, которые до этого отдыхали или занимались другими работами. С понижением температуры частота звуков вентилировщиц уменьшается. Это влечет за собой ослабление активности пчел, занимающихся охлаждением гнезда. Так сопутствующий звук в процессе эволюции общественного образа жизни насекомых обрел чрезвычайно важное значение в системе регуляции терморежима гнезда. Эффективными средствами снижения температуры при перегреве гнезда являются: испарение воды, доставляемой в него пчелами, и уменьшение доли тепла, выделяемого взрослыми особями. Последнее достигается тем, что большая их часть покидает жилище, располагаясь в виде грозди вблизи леткового отверстия. Эта гроздь образуется обычно в дневные часы во второй половине дня и исчезает (пчелы возвращаются в улей) к вечеру. Реакция на охлаждение. Кратковременные небольшие понижения температуры в пчелином гнезде в период высокой активности семьи стимулируют быстрое повышение температуры тела пчел. Например, у пчел, находящихся в зоне с температурой 33° С, температура груди составляет 34—34,5° С. На охлаждение окружающего ее воздуха до 30—32° С температура тела пчел поднимается до 37—38° С со скоростью около 2° С в минуту. Эта температура поддерживается 1—2 мин и затем опускается до 35—35,5° С. Такие скачкообразные изменения температуры груди продолжаются до тех пор, пока окружающий пчелу воздух не прогреется до исходного уровня. Вслед за этим к исходному уровню возвращается и температура груди пчелы (Х. Эш, 1960). При значительных похолоданиях одного увеличения температуры тела пчел недостаточно. Если бы они пользовались только этим, то быстро расходовали свой основной энергетический материал — мед и погибали. Устойчивость пчелиной семьи к длительному и глубокому Охлаждению связана в значительной мере со способностью пчел регулировать тепловую отдачу гнезда через изменение его теплоизоляции. Уже небольшие ночные похолодания в весенне-летний период побуждают пчел, находящихся в различных местах жилища, собираться в зоне сот с расплодом. При этом наиболее плотно они группируются в периферических, более охлаждаемых частях межрамочных пространств, образуя своими телами теплоизоляционную оболочку, способствующую понижению тепловых потерь. Особого внимания заслуживает рассмотрение механизма терморегуляции зимним клубом, представляющим своего рода саморегулирующуюся систему с обратной связью. Образование клуба является реакцией пчел на длительное охлаждение, стимулирующее их собираться в наиболее теплой части жилища. Холод побуждает пчел, образующих периферическую часть клуба, плотно прижиматься к находящимся в более глубоких слоях. Чем дальше от поверхности, тем меньше подвергаются пчелы действию холода. Поэтому плотность клуба от периферии к центру постепенно уменьшается. Неравномерность охлаждения наружной части клуба, связанная с особенностями тепловой защиты жилища и распределением тепла, выделяемого клубом, обусловливает неоднородность плотности в различных его зонах. Наиболее рыхлой бывает верхняя часть клуба, расположенная непосредственно над его тепловым центром. Изменение плотности клуба и соответственно занимаемого им объема является важным механизмом регуляции пчелами тепловых потерь. В частности, уплотнение клуба, предпринимаемое пчелами в ответ на похолодание, влечет за собой снижение тепловых потерь. Теплопотери уменьшаются за счет снижения воздухообмена между внутриклубным пространством и окружающей средой. При этом происходит снижение затрат тепла через излучение, так как уменьшается соотношение между площадью поверхности клуба и его объемом. Своеобразие механизмов терморегуляции у пчел связано в значительной мере со специфичностью работы их терморецепторов. В частности, высокочувствительные холодовые терморецепторы позволяют пчелам контролировать незначительные понижения температуры. Эти рецепторы совместно с тепловыми используются при ориентации по температурному градиенту для того, чтобы находить наиболее теплую зону в жилище. Поведение пчел, пробравшихся в тепловой центр клуба, во многом определяется спецификой работы тепловых рецепторов, являющихся одновременно и рецепторами углекислого газа. Совмещение этих двух функций в одном и том же рецепторе имеет важный биологический смысл. Дело в том, что понижение температуры, стимулирующее уплотнение клуба, ухудшает его аэрацию. Поэтому в нем возрастает концентрация углекислого газа, являющегося продуктом метаболизма пчел. В результате рецептор подвергается здесь одновременному воздействию двух стимулов (углекислоты и высокой температуры), вызывающий однонаправленную реакцию. Возбуждение пчел ведет к повышению температуры в зоне теплового центра. С позиций изложенного становится понятной причина скачкообразного повышения температуры в центре гнезда при резких похолоданиях. Роль расплода. Наличие связи между количеством расплода в семье и температурой в пчелином гнезде послужило основанием для ошибочной интерпретации этого явления. В частности, высказывалось мнение, согласно которому тепло в гнезде пчелиной семьи продуцируется в основном расплодом (О. Мюллер, 1957; И. Плотников, 1961; Т. Жданова, 1963, и др.) и даже регулируется им (И. Плотников, 1961). Рассмотрим специальный опыт И. Плотникова, результаты которого положены им в основу вышеизложенных утверждений. Суть опыта заключалась в следующем. В пустой хорошо утепленный улей с закрытым летком поставили девять сотовых рамок, содержащих в сумме около 22 тыс. ячеек с расплодом. Оказалось, что при колебаниях внешней температуры от 21 до 25° С температура между крайними рамками составляла 30—32° С, а в центральной части — 36° С. В действительности же результаты опыта И. Плотникова лишь подтвержают мысль о том, что посредством теплоизоляции можно сохранить тепло, выделяемое расплодом в процессе метаболизма. Однако здесь уместно отметить, что расплоду наряду с теплом требуется также кислород. Эта потребность расплода удовлетворяется аэрацией гнезда, в которой активно участвуют рабочие пчелы. В процессе аэрации в течение часа из жилища удаляется количество воздуха, многократно превосходящее его объем. Следовательно, результаты опыта И. Плотникова нельзя перенести на условия пребывания расплода в естественных условиях — в семье. Утверждение о том, что расплод активно регулирует тепловой режим, подразумевает следующее: на нагревание он должен реагировать понижением тепловыделения, a на охлаждение — повышением. На самом деле интенсивность метаболизма (для расплода это единственно возможный способ регуляции температуры) с понижением температуры уменьшается, и наоборот. Отсюда при охлаждении количество тепла, выделяемого расплодом, снижается, а при нагревании — возрастает. Доказательством того, что расплод не принимает активного участия в терморегуляции пчелиного гнезда, свидетельствуют результаты опыта (Е. Еськов, 1978), в котором с помощью микротермодатчика регистрировали изменение температуры пчелиного расплода при охлаждении окружающего воздуха. Для этого сотовую рамку, содержащую примерно 4 тыс. ячеек запечатанного расплода, поместили в термостат объемом 0,6 м3. Исходную температуру поддерживали на уровне 34 °С. Затем в течение шести часов ее постепенно понизили до 20 °С. Вместе с понижением температуры воздуха охлаждался расплод. Его температура лишь на 1—2 °С превышала температуру среды. Итак, тепло, выделяемое расплодом, не может играть превалирующей роли в тепловом балансе пчелиного гнезда. Доля его участия в тепловыделении пчелиной семьи бывает, пожалуй, наибольшей в период низкой активности пчел, когда они при кратковременных похолоданиях, собравшись вокруг расплода, образуют своими телами теплоизолирующую оболочку. При этом удерживается тепло, выделяемое пчелами и расплодом. Связь же между стабильностью терморежима в гнезде и наличием в нем расплода говорит о том, что он стимулирует пчел регулировать температуру. Возможно, процесс терморегуляции побуждает какие-то, пока еще неизвестные феромоны, выделяемые живым расплодом. Подтверждением этого предположения может служить тот факт, что пчелый заботятся о живом расплоде, находящемся на любой стадии развития, и удаляют из жилища погибших личинок и куколок.
|
 |
 |

ПОИСК:
|
 |
© PASEKA.SU, 2001–2019
Хаустова Наталья разработка оформления
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paseka.su/ 'Paseka.su: Всё о пчеловодстве'
