 |
Новости |  |
|
Библиотека | |
|
Новые книги | |
|
Ссылки | |
|
Карта сайта | |
|
О нас | |
 |

Сигнал дальностиВременная структура сигнала дальности. В то время, когда танцовщица совершает целенаправленный пробег, в течение которого машет брюшком, она одновременно генерирует звук. Он имеет пульсирующую структуру. Такие звуки пчелы могут генерировать и в тех случаях, когда не исполняют типичных танцев, а находясь в состоянии сильного возбуждения, перемещаются по гнезду. В отличие от этого наблюдаются случаи, когда танцующие пчелы не генерируют звуков. Виляющие движения и генерация звука танцовщицей не взаимосвязанные, а всего лишь совпадающие сигнальные элементы ее поведения. Частота следования импульсов в звуковом сигнале превосходит частоту виляний брюшка в 2,3-2,5 раза. При этом звуковые импульсы не связаны с какой-либо определенной фазой виляющих движений брюшка танцовщицы. Это установили X. Эш (1961) и А. Веннер (1962) одновременной регистрацией звуков и виляющих движений брюшка танцующей пчелы. В опытах X. Эша виляющие движения брюшка танцовщицы регистрировали с помощью индукционного датчика, для чего к дорсальной части брюшка приклеивали миниатюрный магнит. Для регистрации звукового сигнала, использовали микрофон. А. Веннер регистрировал колебания брюшка и звуковые, сигналы, издаваемые танцовщицей, с помощью одного микрофона, к которому был приклеен небольшой лист целлофана. Микрофон держали над танцовщицей так, что она ударяла по целлофану брюшком один раз в течение каждого виляющего цикла. Сигнал дальности представляет собой упорядоченный по амплитуде и частоте следования пакет импульсов. Каждый импульс состоит в среднем из четырех звуковых периодов и длится около 15 мс. Паузы между импульсами имеют примерно такую же величину. Импульсы следуют с частотой около 33 Гц. Длительность импульсов, как и пауз между ними, не связана с расстоянием до цели полета. С ним коррелирует длительность сигнала и количество составляющих его импульсов. Связь между длительностью звукового сигнала танцовщицы и расстоянием до цели полета указывает на способность сигнальщиц измерять пролетаемый ими путь. На чем же основан у них механизм измерения расстояния? Здесь уместно отметить, что этим вопросом интересовались исследователи, изучавшие механизмы, детерминирующие различные динамические позы танцовщицы. Ими, в частности, было обнаружено возрастание длительности виляющей фазы танца в случае увеличения нагрузки летящей пчелы в результате укорачивания крыльев, а также приклеивания к ее груди свинцовой фольги или торможения полета прикрепленными к телу кусочками бумаги. Замечено также, что длительность виляющей фазы танца возрастает и в тех случаях, когда пчела вынуждена летать к источнику пищи в гору или против ветра (X. Херан, Л. Банке, 1952; К. Фриш, X. Херан, М. Линдауер, 1953; X. Херан, М. Линдауер, 1963, и др.) Существование же прямой связи между длительностью виляющей фазы танца и звукового сигнала позволяет предположить, что энергетические затраты танцовщицы на полет к источнику пищи детерминируют соответственно продолжительность звучания сигнальщицы (длительность сигнала дальности). Это предположение было подтверждено опытами, в которых у пчел, посещавших кормушки с 60%-ным раствором сахара на расстоянии 200 м от улья, в то время, когда они, находясь на кормушке, брали сироп, укорачивали одно крыло (Е. Еськов, 1976, 1979). Этим опытом предполагали определить, влияют ли затраты крыловой мускулатуры пчелы на структуру звуков, издаваемых в период танца, и, если это влияние имеет место, то какие затраты (на весь полет или только в одну сторону: от улья к кормушке или обратно) учитываются сигнальщицей при оценке расстояния до источника пищи. Для этого вначале были записаны звуки интактных танцовщиц, а затем звуковые сигналы этих же пчел, летавших с укороченными крыльями только от кормушки до улья. В этот период опыта всех пчел с укороченными крыльями отлавливали и временно исключали из участия в опыте после того, как они, побывав в улье, возвращались на кормушку. Эта мера была предпринята для того, чтобы предотвратить возможность записи звуков пчел-танцовщиц, пролетевших от улья до кормушки и обратно с укороченными крыльями. Опыт завершили записью звуков пчел, летавших от улья до кормушки и обратно с укороченными крыльями. К этому времени выпустили отловленных ранее пчел, а записывать издаваемые ими звуки начали лишь после того, как они стали вновь регулярно посещать кормушку. По окончании опыта отловили пчел с укороченными крыльями в тот момент, когда они, возвратившись после посещения кормушек, входили в летковое отверстие улья. У отловленных пчел взвесили медовые зобики и измерили длину крыльев, определив тем самым, на сколько они были укорочены. Параллельно взвесили медовые зобики интактных пчел, посещавших ту же кормушку, что и подопытные. Взвешивали медовые зобики, заполненные раствором сахара, для того, чтобы определить, не повлияло ли укорачивание крыльев на величину пищевой ноши этих пчел. Оказалось, что укорачивание крыла на 16,8±1,2% (Cυ - 37%) увеличило длительность звукового сигнала и количество составляющих его импульсов. Так, длительность сигнала интактных пчел составляла 609±11 мс (Cυ - 14%); каждый сигнал состоял из 18±0,5 импульсов (Cυ - 21%). Длительность сигнала пчел, летавших с укороченными крыльями только от кормушки до улья, возросла примерно в 1,5 раза, достигнув 906±16 мс (Cυ - 14%). Число импульсов в нем - 25,1±0,5 (Cυ - 15%). Длительность сигнала у пчел, летавших с укороченными крыльями от улья до кормушки и обратно, составляла 864±19 мс (Cυ - 16%) и состоял он из 21,6±0,4 импульса (Cυ - 16%). Взвешиванием медовых зобиков обнаружили, что масса медового зобика у интактных пчел составила 58±2 мг (Cυ - 10,2%), она была такой же у пчел, летавших с укороченными крыльями только от кормушки до улья. У пчелы же, уже из улья вылетавшей с укороченными крыльями, при возвращении в улей медовый зобик весил 41±1 мг (Cυ - 19,4%). Следовательно, увеличение длительности сигнала дальности и количества составляющих его импульсов с ростом затрат энергии сигнальщиц на полете говорит о том, что именно величиной этих затрат руководствуются пчелы, оценивая расстояние до источника пищи. При этом в расчет принимаются лишь затраты энергии на полет от источника пищи до улья. Отметим, что в естественных условиях пчелы в течение одного визита к массиву медоносных растений посещают большое количество цветков. Перелеты от одного цветка к другому сопряжены с определенными затратами энергии крыловой мускулатуры. Немало этой энергии тратят пчелы и на формирование пыльцевой ноши. Пчела формирует эти ноши на третьей паре ног, находясь в воздухе. Для выявления возможного влияния затрат энергии при формировании пыльцевой ноши на структуру сигнала дальности был проведен сравнительный анализ звуков, издаваемых одними и теми же пчелами, приносившими попеременно углеводную пищу или пыльцу. В качестве углеводной пищи давали 60%-ный раствор сахара. Предлагаемая подопытным пчелам пыльца растений представляла собой высушенные и измельченные до пылеобразного состояния пыльцевые ноши (обножки), отобранные у других пчел. Взятие пчелами пыльцы и раствора сахара было сопряжено с различными затратами энергии крыловой мускулатурой. Во время заполнения медового зобика раствором сахара пчелы сидели на кормушке, а при формировании пыльцевой ноши - многократно взлетали. Время, в течение которого пчела, формируя пыльцевую ношу, находилась в воздухе, составляло 112±9 с (Cυ - 18%), что значительно превосходило время на обратный полет, который длился как с той, так и с другой ношей не более 6Чёс. Это, тем не менее, не отразилось на длительности и числе импульсов в сигнале дальности (табл. 5).  Таблица 5. Временная структура сигнала дальности пчел, приносящих в улей раствор сахара или цветочную пыльцу Следовательно, энергетические затраты на формирование пыльцевой ноши, а также перелеты пчелы от одного цветка к другому при сборе нектара не принимаются во внимание сигнальщицей. Начальным пунктом отсчета пролетаемого пути служит местонахождение последнего из посещаемых пчелой цветка. При этом структура сигнала не зависит от того, собирала ли пчела нектара и пыльцу порознь, вместе или только одно из этих веществ. Наряду со значительными энергетическими затратами на полет и сбор пищи, немало энергии расходуют пчелы на перемещение внутри улья - от летка в гнездо и к летковому отверстию. Энергетические затраты на преодоление равного отрезка пути летящей и бегущей пчелой различаются в десятки раз. Например, по данным Е. Сшользе и соавт. (1964), пчела, пробежавшая путь длиной 3 м, потребляет такое же количество сахара, как при полете на расстояние около 60 м. Чтобы определить, влияют ли затраты энергии ножной мускулатурой сигнальщиц на структуру их сигналов, провели специальные опыты, в которых вылетающих и возвращающихся в улей пчел вынуждали проходить по затемненному лабиринту разной длины. Лабиринт, построенный в виде узкого коридора шириной 3 и высотой 5 см, был прикреплен к летковому отверстию наблюдательного улья, находившегося в светонепроницаемой камере. Небольшая группа меченых пчел этого улья посещала кормушку с 60%-ным раствором сахара на расстоянии 200 м от улья. Сопоставление результатов осциллографического анализа сигналов танцовщиц, проходивших по лабиринту путь длиной 0,1 и 3,5 м, свидетельствует о том, что затраты энергии ножной мускулатуры не отражаются на длительности сигнала и числе составляющих его импульсов (табл. 6). Отсюда измерение расстояния до цели полета ведется сигнальщицей исключительно по затратам энергии крыловой мускулатуры. Конечный пункт отсчета расстояния - место посадки прилетевшей пчелы летковое отверстие улья).  Таблица 6. Временная структура сигналов дальности у пчел, живущих в улье с легковыми лабиринтами разной длины (по Е. Еськову, 1973) Увеличение пешего пути вызвало в описываемом опыте лишь уменьшение примерно на 30% длительности пауз между сигналами (табл. 6). Вместе с тем было замечено, что пчелы не безразличны к длине пути, который они вынуждены проходить от гнезда до леткового отверстия. Выяснилось, что пчел не удовлетворяет жилище, которое связано с внешней средой длинными ходами. Такие жилища они покидают. Критическая длина леткового лабиринта для пчел серой горной кавказской расы составляет 2,5-3 м. При такой его длине они начинают поиск нового жилища и, если летковый лабиринт не укоротить, то вскоре улетают, бросив гнездо, даже если в нем есть расплод. На временную структуру сигнала дальности не влияет температура внешней среды. Во всяком случае не обнаружено изменение сигнала при изменении внешней температуры с, 17° до 31° С. Однако значительные колебания внутриульевой температуры отражаются на временной структуре сигнала. Это выявлено в опытах на семьях, содержащихся в специальной терморегулируемой камере. Оказывается, внутриульевая температура влияет на длительность сигнала и частоту следования составляющих его импульсов. Понижение температуры с 27-37° до 16-17° С уменьшает частоту следования импульсов на 3,7-5,6 Гц и число звуковых периодов в нем на 11,5%. Примерно на столько же увеличивается длительность всего сигнала. Для выявления влияния числа пчел в семье (силы семьи) на структуру издаваемых ими сигналов дальности были проведены специальные исследования на семьях разной величины. Исходно использованы семьи средней величины, число особей в которых уменьшали по ходу опытов разными способами. В одном из таких опытов исходно использовали семью, в которой насчитывалось примерно 25 тыс. рабочих особей. Группа меченых пчел этой семьи, жившей в 12-рамочном улье, посещала кормушку с раствором сахара на расстоянии 20 м от улья. Звуки, издаваемые этими пчелами, регистрировали в течение нескольких дней. Затем на место 12-рамочного поставили пустой однорамочный улей, в который перенесли одну из рамок от подопытной семьи вместе с находившимися на ней пчелами, маткой и разновозрастным расплодом. Заменяли улей в то время, когда пчелы активно посещали кормушку. Поэтому большая часть меченых фуражиров оказалась в однорамочном улье. Звуки, издаваемые мечеными пчелами, посещавшими кормушку, записывали 14 дней с момента организации новой семьи. Во время проведения этих записей внешняя температура среды изменялась с 26° до 29° С. Анализ временной структуры сигналов пчел, живущих в сравнительно большом 12-рамочном и маленьком однорамочном ульях, свидетельствует о том, что величина семьи не влияет на характеристики звуков, издаваемых фуражирами (табл. 7).  Таблица 7. Временная структура сигналов дальности у пчел, живущих в семьях, разной величины (по Е. Еськову, 1969) Влияние возраста сигнальщиц на структуру издаваемых ими звуков изучали на пчелах разных рас, отличающихся по окраске тела. В частности, однодневных пчел итальянской расы (с желтыми кольцами на брюшке) помещали в семью краинской расы (с серым брюшком), содержащуюся в однорамочном наблюдательном улье. Как только пчелы итальянской расы приступили к выполнению роли фуражиров (с 14-17-дневного возраста), их приучили посещать кормушку с раствором сахара на расстоянии 200 м от улья, на которую они летали до последних дней жизни. В этом опыте не обнаружили влияния возраста сигнальщиц на спектральную структуру, длительность и число импульсов в издаваемых ими звуках. Возраст сигнальщиц отражается лишь на временной структуре импульсов, видоизменяющихся по мере старения пчел. Эти изменения выражаются прежде всего в увеличении коэффициента вариации числа звуковых периодов, образующих импульс. В сигналах очень старых пчел импульсы не всегда разделены четкими паузами. Длительность и число импульсов изменяются неодинаково в сигналах дальности пчел разных рас в связи с увеличением расстояния от улья до цели полета (Е. Еськов, 1968, 1969, 1970; Н. Лопатина, 1971; Н. Лопатина, В. Пономаренко, 1972). Например, с увеличением расстояния до цели полета с 30 до 200 м число импульсов в сигналах пчел итальянской расы возрастает в среднем на 12, серой горной кавказской - на 11, а краинской - всего на 3. Соответственно увеличение длительности сигнала при тех же изменениях расстояния у пчел итальянской и серой горной кавказской рас больше, чем у пчел краинской расы (рис. 11). 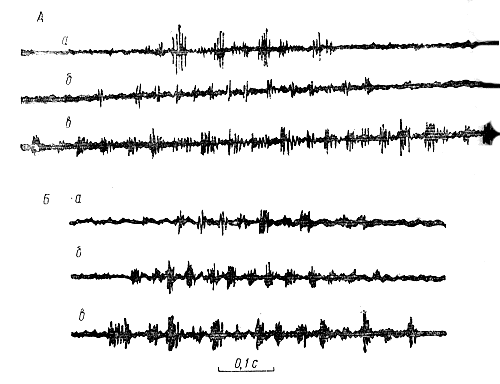 Рис. 11. Осциллограммы сигналов дальности пчел итальянской (Л) и краинской (Б) рас. оповещающих членов семьи об источниках пищи, расположенных на расстоянии 30 (а), 100 (б) и 200 м (в) от улья (по Е. Еськову, 1969) Расовая специфичность сигналов дальности имеет наследственную природу. Это показано в опытах с пчелами, воспитанными и живущими с рождения в семьях других рас (Е. Еськов, 1971, 1972). Исследования проводили на пчелах краинской и итальянской рас. Эти расы использовали потому, что у них существенно различаются по длительности сигналы при посещении равноудаленных источников пищи, особенно на расстоянии 200 м от улья. Кроме того, отличия в окраске пчел этих рас давало возможность различать их, не прибегая к мечению. Во избежание контактов подопытных пчел с особями своей расы запечатанный расплод помещали в термостат за 1-3 дня до выхода взрослых пчел, которых затем подсаживали семье другой расы. В другой серии опытов подопытные пчелы не только жили с момента рождения в семье другой расы, но и были выращены в ней. Для этого соты с яйцами из семьи итальянской расы переносили в семью краинской расы, живущую в обычном 12-рамочном улье. Примерно за сутки до появления пчел эти соты переносили в наблюдательные ульи, в которых жили семьи краинской расы. Таким образом, пчелы итальянской расы не имели контактов с особями своей семьи на протяжении всего периода постэмбрионального развития. Во многих из этих опытов подсаженные пчелы и пчелы-хозяева начиная с первого вылета в течение длительного времени посещали одни и те же кормушки с раствором сахара, лишь изредка переключаясь на другие, естественные источники пищи. Сигналы дальности тех и других пчел регистрировали в одно и то же время по возвращении их с кормушки. Опыты показали, что воспитание пчел с момента рождения в семье другой расы не влияет на структуру сигналов дальности. Например, сигналы пчел краинской расы, помещенных с первого дня жизни в семью итальянской расы и, следовательно, не имевших в процессе онтогенеза контакта с особями своей расы, не имели достоверных различий с сигналами индивидов той же расы, постоянно живших в собственной семье, и наоборот (табл. 8). Аналогичный результат получен в опытах с пчелами итальянской расы, выращенными из яиц семьями краинской расы.  Таблица 8. Сигналы дальности у пчел, воспитанных в своей семье и семье другой расы (по Е. Еськову, 1971) Спектральная структура сигнала дальности. Основная энергия сигнала дальности лежит в области 160-600 Гц с двумя максимумами на частотах 200-350 и 400-600 Гц. Первый из этих диапазонов интенсивнее второго на 10-20 дБ. Спад интенсивности сигнала на область высоких частот составляет в среднем 11 Дб на октаву. Диапазон максимальных по интенсивности составляющих отличается наибольшей стабильностью. Спектральная структура сигнала не зависит от указываемого сигнальщицами расстояния до цели полета. Частотное положение спектрального максимума в сигналах, издаваемых на относительно невысоком фоне естественных звуков пчелиной семьи, изменяется от импульса к импульсу в небольших пределах (рис. 12). Значительные изменения в спектре сигнала происходят при больших изменениях интенсивности и спектральной структуры звукового фона улья или посторонних звуковых помехах. 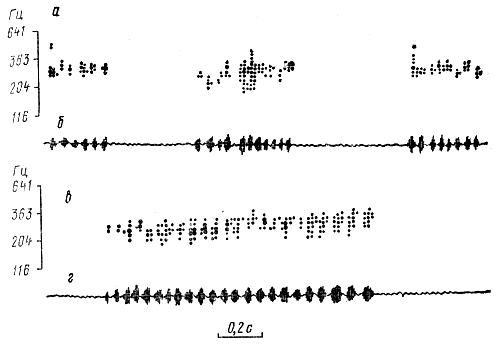 Рис. 12. Динамические спектры (а, в) и осциллограммы (б, г) сигналов дальности при различном расстоянии до цели полета; а, б - 30 м; в, г - 200 м (по Е. Еськову, 1979) Влияние звукового фона семьи на структуру сигналов дальности было прослежено в процессе анализа одновременно записанных внутри улья сигналов танцовщиц и звуков, на фоне которых осуществляется эта сигнализация. Для этого звуки фуражиров, посещавших кормушки с раствором сахара на расстоянии 200 м от улья, записывали на магнитной ленте. Микрофон подносили к танцовщице, располагая его на расстоянии 8-12 мм над поверхностью ее крыльев. В результате вместе с сигналами танцовщиц на ленте записывались другие звуки, издаваемые обитателями улья. Анализ записанных звуков проводили следующим образом: участки магнитной ленты с записью сигналов танцовщиц и фоном, записанным в паузах между этими сигналами, порознь склеивали в кольца и анализировали. Для анализа использовали анализаторы частотно-амплитудного и динамического спектров. В результате установлено, что интенсивность фоновых составляющих на расстоянии 8-12 мм от сигнальщиц обычно значительно ниже интенсивности составляющих сигнала. В семьях, находящихся в спокойном состоянии, интенсивность составляющих сигнала в диапазоне 200-350 Гц превосходит соответствующую фоновую область на 15-18 дБ (Cυ - 35%). Диапазон интенсивности высокочастотных составляющих сигнала выше совпадающих с ним фоновых на 3-6 дБ (Cυ - 34%). В отдельных случаях, когда обитатели улья сильно возбуждены, интенсивность фона резко возрастает. В это время звуковой фон улья может достигать, а иногда даже превосходить по интенсивности составляющие сигнала дальности. Частотное положение интенсивных составляющих сигнала определяется спектральной структурой фона и зависит от различий интенсивностей сигнала и фона. При большом различии интенсивностей сигнала и фона частотное положение сигнального максимума интенсивности совпадает с областью максимально интенсивных составляющих фона. В соответствии со смещением диапазона интенсивных спектральных составляющих фона, обусловленным изменением состояния пчелиной семьи, смещается диапазон интенсивных составляющих сигнала. Диапазоны совпадающих по частоте интенсивных спектральных составляющих сигнала и фона уменьшается по мере увеличения интенсивности фона и связанного с этим уменьшением различий интенсивностей сигнала и фона. При различии интенсивностей сигнала и фона не менее 17 дБ диапазоны интенсивных сигнальных и фоновых составляющих практически совпадают. С уменьшением этого различия до 13-16 дБ спектр интенсивных составляющих сигнала смещается по отношению к частотному положению интенсивных составляющих фона в сторону высоких частот. Диапазон смещения достигает примерно 50 Гц. Это выражается в основном в более пологом спаде высокочастотных составляющих сигнала по сравнению с фоновыми. Спад интенсивности высокочастотной части в спектре сигнала составляет примерно 3 дБ на октаву, а в фоне - около 10 дБ. В тех случаях, когда интенсивность фона приближается к интенсивности сигнала, например перед слетом пчелиной семьи, диапазон интенсивных составляющих в звуках танцовщиц значительно расширяется, что происходит в значительной мере за счет изменения частотного положения его энергетического максимума от импульса к импульсу. При этом существенно возрастает диапазон интенсивных составляющих сигнала, не совпадающих по частотному положению с интенсивными составляющими фона. В результате большая часть энергии сигнала сосредоточивается в области, свободной от интенсивных фоновых составляющих. Наличие связи между спектральной структурой сигналов танцовщиц и звуков, на фоне которых осуществляется эта сигнализация, подтверждено в опытах с генерацией искусственных помех. Оказывается, изменение интенсивности и спектральной структуры посторонних для пчел звуков, как и естественного звукового фона улья, влияет на частотное положение интенсивных составляющих сигнала и ширину его спектра. Кодирование танцовщицей информации о расстоянии до указываемой цели. Первые попытки расшифровать код пчелиного танца были предприняты в конце 50-х годов 20 века. Для этого вначале применили модели танцовщиц, например, деревянные цилиндры величиной с пчелу или мертвых пчел (В. Штехе, 1957; X. Эш, 1964, и др.). Ими имитировали движения танцовщиц. Однако это не имело успеха, хотя пчелы проявляли интерес к модели танцовщиц. После того как было обнаружено, что танцовщицы генерируют звуки, их стали воспроизводить с помощью громкоговорителей, расположенных рядом с ульем. X. Эш (1964), а затем Н. Лопатина, Е. Чеснокова (1967) и И. Левченко (1976) включали звук в период, когда модель имитировала виляющие движения танцовщицы. Но и это не стимулировало целенаправленного вылета новичков. Такой результат объясняется тем, что условия этих опытов слишком далеки от естественного и не удовлетворяют многим из условий, необходимых для обеспечения коммуникационного процесса между танцовщицей и мобилизуемыми ею пчелами. Предпосылкой для предположения о том, что информация о расстоянии кодируется в длительности акустического сигнала танцовщицы, послужило сопоставление результатов работ В. Штехе (1954) и Р. Боха (1957) на смешанных семьях, в которых пчелы итальянской расы неправильно ориентировались по танцам пчел краинской расы и, наоборот, с обнаруженными позже (Е. Еськов, 1968) различиями в длительности звуковых сигналов пчел этих рас, посещающих кормушки на одинаковом расстоянии от улья. В частности, в опытах В. Штехе и Р. Боха было замечено, что большинство пчел итальянской расы, мобилизованных фуражирами краинской расы, посещавшими кормушки на расстоянии 160 м от улья, прилетали к кормушкам, расположенным в том же направлении на расстоянии 120 м. В отличие от этого пчелы краинской расы, мобилизованные фуражирами итальянской расы, искали источник пищи дальше того места, где находилась кормушка. Результаты этих опытов объясняются различиями в длительности звуковых сигналов дальности пчел краинскойи итальянской расы, оповещающих о равноудаленном источнике пищи (Е. Еськов, 1968). Действительно, длительность звуковых сигналов дальности пчел краинской расы, посещающих источники пищи на расстоянии 120 и 160 м от улья, соответственно на 60 и 150 мс меньше длительности сигналов пчел итальянской расы. Информационная роль звуков танцовщиц как сигналов, используемых для оповещения о расстоянии до цели; полета, была доказана путем увеличения их длительности (Е. Еськов, 1971). При постановке опыта исходили из того, что если информация о расстоянии кодируется в длительности сигнала, то дополнительным воспроизведением его определенной части без нарушения естественного контакта танцовщицы со свитой можно увеличить дистанцию, направленного полета новичков. Длительность сигнала танцовщицы увеличивали с помощью специальной системы с запоминающим устройством, содержащим микрофон, усилитель, магнитофон с двумя раздельными блоками (один для записи, другой для воспроизведения) и звуковые излучатели. Зонд микрофона помещали над танцовщицей, а входные отверстия излучателей - над окружающими ее (мобилизуемыми ею) членами свиты. Влияние увеличения длительности сигнала на оценку пчелами свиты расстояния до цели полета определяли по числу новичков, прилетавших к трем кормушкам, которые расположили по линии лета фуражиров. Расстояние до первой кормушки 100 м, до второй (ее посещали фуражиры) - 150 м, до третьей - 200 м. Сигнал увеличивали на 200 мс, что соответствовало его длительности при посещении фуражирами цели полета на расстоянии 200 м от улья, В результате увеличения длительности сигнала та часть новичков, которая слышала этот сигнал, прилетала к кормушке, расположенной на расстоянии 200 м от улья. Это указывает на то, что в длительности пульсирующего звукового сигнала танцовщиц кодируется информация о расстоянии до цели полета. Пользуясь звуковыми сигналами танцовщиц, окружающие ее пчелы могут получать, по-видимому, информацию и о направлении к цели полета. Основанием для этого служит, во-первых, целенаправленное перемещение - пчелы-сигнальщицы и, во-вторых, наличие у пчел тонкого бинаурального слуха. Благодаря этому мобилизуемые пчелы в состоянии контролировать перемещение танцовщицы. При этом собственное положение на вертикальной поверхности сот в темном жилище они определяют с помощью хорошо развитых гравитационных рецепторов. Для передачи сигналов дальности пчелы пользуются воздушной линией связи. Небольшая интенсивность этих сигналов и значительное поглощение звуковой энергии воздушной средой ограничивает радиус их действия несколькими миллиметрами. Но именно в этой зоне и находятся пчелы свиты, мобилизуемые сигнальщицами. Слабая интенсивность сигналов дальности обеспечивает возможность одновременной передачи сообщений несколькими сигнальщицами, находящимися на одном и том же соте. Это важно, так как эффективность мобилизации во многом зависит от числа сигнальщиц, которые при большой интенсивности сигналов мешали бы друг другу. Более высокая интенсивность сигналов нецелесообразна еще и потому, что при передаче других не акустических сигналов, входящих в систему мобилизации, новички также должны находиться вблизи сигнальщицы. Следовательно, слабая интенсивность сигналов дальности необходима и достаточна для передачи их по воздушной линии связи, Сигнал, останавливающий танцовщицу. Пчелы, проявляющие интерес к танцовщице, получают от нее информацию о качестве пищи в процессе пищевых контактов. Обычно танцовщица перед танцами и в паузах между их циклами сама предлагает другим пчелам попробовать принесенную пищу. При необходимости пчелы, следовавшие за танцовщицей в течение нескольких танцевальных циклов, могут останавливать ее акустическими сигналами для взятия пробы пищи. Сигнал, останавливающий танцовщицу, содержит всего один импульс длительностью 185±90 мс. Начало сигнала длительностью 25±10 мс характеризуется наличием двух диапазонов интенсивных составляющих - на частотах 200-400 и 500-580 Гц. Первые интенсивнее вторых примерно на 10 дБ. Частотное положение первого спектрального максимума энергии плавно изменяется с 200-330 Гц в начале этого участка до 250-400 Гц к его концу. В дальнейшем интенсивные составляющие сигнала сохраняются только в этом диапазоне частот. Важно отметить, что танцовщица останавливается лишь в том случае, если сигнал по структуре соответствует описанному. Сигналы, имеющие аналогичную амплитудно-временную структуру, но отличающиеся по спектру, не останавливают танцовщицу (Е. Еськов, 1972). Роль звуков как сигналов остановки танцовщиц подтверждена опытами X. Эша (1964) с использованием деревянной модели пчелы, имитирующей движения танцовщицы. Двигающаяся модель привлекала внимание окружающих ее пчел, которые издавали время от времени звуковые сигналы, используемые для остановки танцовщицы. Если модель после этого продолжали двигать, пчелы набрасывались на нее, чего не наблюдалось в тех случаях, когда по сигналу остановки танцовщицы модель немедленно останавливали. Ольфакторная ориентация и сигнализация. В процессе поиска источника пищи пчелы-новички руководствуются преимущественно информацией, заключенной в танцах мобилизовавших их фуражиров. Насколько строго придерживаются пчелы-новички этой информации, говорят опыты К. Фриша (1955), в которых по линии лёта от улья к кормушке с раствором сахара, имевшего слабый запах лаванды, лежали пластины, пропитанные тем же пахучим веществом. В разных вариантах опыта кормушка находилась на расстоянии 750 или 2000 м от улья. Ароматизированные пластины лежали на расстоянии 75, 200, 400, 700, 800, 1000, 1500, 2000 и 2500 м от улья. Несмотря на это большинство пчел-новичков, мобилизованных танцами фуражиров, прилетало к кормушкам, не обращая внимания на пахучие пластины. Вместе с тем залах пищи, приносимый фуражирами, принимается во внимание новичками. Прилетев к указанному месту, они посещают в первую очередь те растения, запах которых был принесен мобилизовавшими их пчелами. Это доказал в своих опытах К. Фриш (1956, 1966). Он приучал группу фуражиров посещать букеты альпийских фиалок, обрызганных раствором сахара. После того как фуражиры, возвратившись в улей, начинали танцевать, эти букеты убирали, а на их место ставили такие же, но без сахарной приманки; рядом выставляли букеты флоксов, обрызганные раствором сахара. Прилетевшие новички долго обследовали альпийские фиалки, не обращая внимания на букеты флоксов. Аналогичный результат получен и в обратном варианте опыта, когда фуражиры приносили раствор сахара, собранный на флоксах. В этом случае новички после замены букетов искали приманку на флоксах, в то время как рядом находились обрызганные сахаром букеты фиалок. Информацию о запахе пищи новички получают в процессе тактильных контактов и трофолаксиса с фуражирами. Запах источника пищи, посещаемого фуражирами, может удерживаться на их теле в течение одного-двух дней (М. Линдауер, 1971). Недооценка этого свойства покровов пчелиного тела послужила, в частности, одной из причин неверного объяснения А. Веннером, Д. Джонсоном, П. Велсом и Ф. Роучифом (1966, 1967, 1969) результатов поставленных ими опытов, которые будто бы опровергают теорию К. Фриша о том, что сообщение о цели полета передается фуражирами в процессе танцев. Это «опровержение» привлекло внимание многих исследователей, вовлеченных в полемику о роли танцев. Полемические статьи на эту тему продолжают появляться в печати. Поэтому в сути вопросов, поднятых А. Веннером и соавт., следует разобраться. А. Веннер и соавторы приписывают решающую роль в мобилизации новичков запаху пищи. В качестве одного из главных аргументов они приводят результаты опытов с тремя кормушками, находившимися примерно на равном расстоянии от улья, но в разных от него направлениях: первая - на юго-западе, вторая - на западе, третья - на северо-западе. Две группы фуражиров в течение одного-двух дней посещали первую и третью кормушки с раствором сахара, ароматизированного запахом клевера (каждая группа фуражиров посещала свою кормушку). В этот период в улье накапливался раствор сахара, ароматизированный клевером. В последующие один-два дня во вторую кормушку наливали раствор сахара, ароматизированный клевером, а в первую и третью - неароматизированный. Меченые фуражиры продолжали посещать первую и третью кормушки. Всех мобилизованных ими новичков отлавливали в первый же визит к каждой из трех кормушек. В результате на первой кормушке отловили 23, на второй - 183, на третьей - 31 новичка. На основании этого А. Веннер и соавторы пришли к выводу о том, что новички возбуждались танцами, но в поисках пищи руководствовались исключительно запахом клевера, который накапливался в улье в период дрессировки фуражиров. Ясность в трактовку результатов описанных опытов внес М. Линдауер (1971), который, в частности, доказал, что новички не принимают во внимание запах пищи, накапливаемой в улье. Для этого М. Линдауер использовал также три кормушки, расположенные, как в опытах А. Веннера. Но в опытах М. Линдауера в первой и третьей кормушках постоянно находился неароматизированный раствор сахара. Запах приманки второй кормушки, ароматизированной чебрецом, искусственно вводили в улей, помещая в него фильтровальную бумагу, пропитанную этим пахучим веществом. За время опыта на всех трех кормушках было отловлено в сумме 140 новичков, в том числе 83 - на первой кормушке, 52 - на третьей и только 5 - на второй. Так был опровергнут один из ошибочных выводов А. Веннера о роли запаха пищи, накапливаемой в улье. Большее же количество новичков, прилетевших в опытах этих исследователей на вторую кормушку, видимо, связано с тем, что новички пользовались запахом клевера, который, как отмечалось выше, мог сохраняться на теле фуражиров. Серьезная ошибка в объяснении А. Веннером и соавторами результатов своих опытов связана также и с тем, что они не учли способности пчел-новичков усреднять информацию о направлении к цели полета, получаемую от разных сигнальщиц, что было замечено Д. Отто еще в 1959 г. Это подтверждается в другой серии опытов, М. Линдауера, проведенных им на тех же трех кормушках. В этих опытах первую и третью кормушки по-прежнему посещали две разные группы фуражиров, но в отличие от предыдущего опыта в этом - все три кормушки были постоянно наполнены раствором сахара, ароматизированного чебрецом. Подопытные пчелы жили в наблюдательном улье, что позволяло следить за поведением новичков, мобилизуемых фуражирами. Оказалось, что часть новичков до вылета из улья успевала побывать в свитах фуражиров, посещающих и первую, и третью кормушки. В результате они усредняли информацию о месте нахождения источника пищи, полученную от разных танцовщиц, посещающих разные кормушки, что подтвердили результаты отлова новичков. Так, на первой кормушке отловили 146 новичков, на второй - 91, на третьей - 203 пчелы. Показателен также результат, полученный после того, как фуражиров, посещавших первую кормушку, исключили из опыта. В этом случае за 30 мин на первую и вторую кормушки прилетело всего по одному новичку, а на третью - 55. Следовательно, мобилизуемые фуражирами пчелы, вылетая из улья, знали, где находится источник пищи. Если бы они ориентировались только по запаху, как это утверждает А. Веннер, то они прилетали бы в равном количестве ко всем трем кормушкам. Все изложенное выше позволяет заключить, что одного запаха пищи недостаточно для мобилизации новичков. Но это не распространяется на пчел, ранее посещавших ароматизированный источник пищи. Для возобновления их полетов достаточно ввести в улей запах знакомого им источника пищи (Н. Смарагдова, 1933; К. Фриш, 1943; А. Губин, 1947; К. Риббанс, 1954, 1955, 1956). Более того, отдельные группы пчел одной и той же семьи, посещавшие разные источники пищи, могут быть избирательно мобилизованы на любой из них введением в улей соответствующих запахов (С. Розов, Н. Сахаров, 1937; К. Риббанс, 1954). Запах корма имеет важное значение для его обнаружения в то время, когда пчела подлетает к месту, указанному танцовщицей. Подлетев, например, к кормушке с раствором сахара, новичок, прежде чем сесть на нее, некоторое время летает перед ней по зигзагообразной траектории. Время этого полета сокращается в том случае, когда раствор сахара в кормушке ароматизирован. О влиянии запаха корма на эффективность мобилизации новичков говорит следующий опыт, проведенный на семье, содержавшей около 30 тыс. рабочих особей. Две группы пчел этой семьи приучили посещать кормушки с раствором сахара, расположенные на расстоянии 300 м от улья. Кормушки находились в разных направлениях от него под углом 90°. Исходно в кормушках находился неароматизированный 50%-ный раствор сахара, на который приучали фуражиров. По 20 наиболее активных из них, летавших к той и другой кормушкам, были помечены краской. Затем раствор сахара в кормушках заменили на 10%-ный. В это время отловили всех немеченых пчел. После того как их прилет прекратился (меченые фуражиры не прекращали летать на кормушки), кормушки вновь заполнили 50%-ным раствором сахара: в одной из них он был неароматизированным, в другой - ароматизирован анисовым маслом. Повышение концентрации раствора сахара стимулировало процесс мобилизации. К кормушкам стали прилетать новички, которых отлавливали и исключали из опыта. За 45 мин на кормушке с ароматизированным кормом быдо отловлено 248 пчел. На кормушку с неароматизированным раствором сахара за это же время прилетело 176 новичков. Следовательно, запах повысил эффективность кор-мособирательной деятельности пчел, что выразилось в увеличении числа их прилета за равное время примерно на 40%, Аналогичный результат был получен после того, как кормушки с ароматизированным и неароматизированным растворами поменяли местами. Перед этим вначале отловили немеченых пчел. Для чего, Как и ранее, кормушки наполняли 10%-ным неароматизированным раствором сахара. Пчелы, посещавшие определенный источник пищи, запоминают его местоположение по визуальным ориентирам. Но и для них запах корма имеет немаловажное значение. Об этом говорит, например, реакция пчел, длительное время посещающих кормушку с раствором сахара, на его ароматизацию или изменение запаха. Пчелы, находящиеся на кормушке и занимающиеся заполнением медовых зобиков, взлетают, если корм или только кормушку ароматизировать. Взлетев, пчелы кружатся 1-2 мин, после чего их поведение нормализуется: они садятся на кормушку, берут корм, по возвращении в улей ведут себя так же, как до ароматизации корма или изменения его запаха. Пчелы, подлетевшие к кормушке после ароматизации (переароматизации) корма, дольше обычного кружатся над кормушкой. В последующие визиты они ведут себя, как и до изменения запаха корма. Важное значение приобретает запах как ориентир в тех случаях, когда местоположение источника корма меняется, что бывает в естественных условиях, которые легко смоделировать, манипулируя кормушками. Например, в одном из опытов группу пчел, которых пометили краской, приучили посещать кормушку с 50%-ным раствором сахара, ароматизированного гераниевым маслом. Для оценки роли этого пахучего ориентира кормушку, посещаемую пчелами после отлова новичков, который проводили, как и в ранее описанных опытах, убирали. Вместо нее ставили две другие. Они были расположены с двух сторон от того места, где находилась приманочная кормушка. Расстояние между ними составляло 50 см. В одной из этих кормушек был 50%-ный раствор сахара, ароматизированный гераниевым маслом, в другой - анисовым маслом. В результате за 30 мин, в течение которых отлавливали всех пчел, садящихся на кормушки, к первой из них прилетело 54 меченых и 37 немеченых особей. Другую кормушку с 50%-ным раствором сахара, ароматизированным анисовым маслом, посетило за то же время 12 меченых пчел и три новичка. Ароматические вещества, использованные в описанном опыте, не обладали сами по себе привлекающим действием на пчел. На это указывает результат другого опыта, проведенного по аналогичной методике, с той лишь разницей, что пчел приучали к запаху анисового масла. Кормушку с ароматом анисового масла посетило в течение 30 мин 57 меченых и 23 немеченых пчелы. На кормушке с гераниевым маслом отловили 14 меченых и одну немеченую пчелу. Итак, запах пищи используется в качестве локального ориентира как новичками, так и особями, занимающимися в течение длительного времени ее доставкой в жилище. В тех же случаях, когда пища не имеет запаха для пчел, что маловероятно в естественных условиях их обитания, пчелы не лишаются возможности использования ольфакторной ориентации. Оказывается, медоносные пчелы подобно другим общественным насекомым таким, как муравьи, термиты и тригоны, могут метить кормовой участок собственными пахучими выделениями. Пахучие вещества могут выделять железы лапок ног, ротового аппарата и насонова органа. Они, абсорбируясь на кормовом участке, выполняют роль пахучих меток. Они используются как новичками, так и мобилизовавшими их пчелами. Об этом говорит следующий опыт. Группу пчел приучили посещать кормушку с неароматизированным 50%-ным раствором сахара. Кормушка находилась на расстоянии 200 м от улья. Прилетающих пчел метили краской. В течение часа пометили 50 пчел. После чего эту приманочную кормушку переместили на 30 см от того места, где она находилась. На таком же расстоянии от него, но с другой стороны поставили аналогичную кормушку с таким же неароматизированным раствором сахара. Но ни с этой кормушкой, ни с раствором сахара, которым ее наполняли, пчелы ранее не контактировали. В течение 30 мин на первой кормушке отловили 46 меченых и 11 немеченых пчел. На второй Ч только три меченых пчелы. Новички совсем не подлетали к ней.
|
 |
 |

ПОИСК:
|
 |
© PASEKA.SU, 2001–2019
Хаустова Наталья разработка оформления
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paseka.su/ 'Paseka.su: Всё о пчеловодстве'
