 |
Новости |  |
|
Библиотека | |
|
Новые книги | |
|
Ссылки | |
|
Карта сайта | |
|
О нас | |
 |

Наследственность и изменчивостьЯвления изменчивости и наследственности свойственны всем живым организмам, и этим кругом вопросов занимается биологическая наука генетика, которая изучает законы, определяющие сходство и различия между особями, родственными друг другу по происхождению. Все живые организмы состоят из клеток. Клетки в большинстве случаев имеют очень маленькие размеры и их можно видеть лишь под микроскопом. Высшие организмы состоят из множества клеток, в то время как бактерии и многие простые организмы имеют одну единственную клетку. В каждой клетке имеется ядро. Ядро, в свою очередь, содержит структурные образования, которые хорошо видны под микроскопом на некоторых стадиях жизни клетки. Из-за способности интенсивно окрашиваться определенными красителями их назвали хромосомами (хромосома от греческого chroma — цвет, — краска, + soma — тело, означает окрашенное тело). Хромосомы являются материальными носителями наследственности и именно они обеспечивают сходство между родителями и потомками. Передаваясь из поколения в поколение, хромосомы как бы несут завещание предков: какой формы, величины и окраски иметь тело, как расти, как плодоносить и т. д. И потомки точно выполняют этот приказ, написанный с помощью хромосом. В клетках тела животных хромосомы всегда парны (Исключение составляет лишь одна пара хромосом, называемых половыми, которые у животных определяют пол особи ), т. е. в любой клетке имеется по две одинаковые хромосомы каждого сорта. Такие хромосомы получили название гомологичных (одинаковых) хромосом. Одну половину хромосом потомство получает от матери, а вторую (гомологичную) — от отца. Хромосомы родителей своим детям передаются через гаметы (яйцеклетки и сперматозоиды), которые вырабатываются половыми клетками. В половых клетках обычное число хромосом, характерное для каждого вида, уменьшается вдвое, а при оплодотворении яйцеклетки сперматозоидом вновь восстанавливается обычное (двойное) число хромосом. Одинарный набор хромосом (в гаметах) носит название гаплоидного числа, а двойной набор хромосом - диплоидного числа. Величина, форма и число хромосом в ядре у разных видов животных и растений неодинаковы. Например, каждая клетка человека имеет 46 хромосом, в то время как у медоносной пчелы (матки и рабочие особи) имеется в ядрах клеток по 32 хромосомы. Однако у медоносных пчел имеются существенные отличия от вышеописанного механизма передачи наследственности родителей своим потомкам и определение пола особей. Если у всех сельскохозяйственных животных пол особи определяется определенными половыми хромосомами, то у медоносных пчел пол особи определяется по другому: при оплодотворении яиц из них образуются самки (матки и пчелы), а из неоплодотворенной яйцеклетки матки развиваются самцы (трутни). Таким образом, женские особи семьи являются диплоидными организмами (имеют 32 хромосомы), а мужские особи (трутни) — гаплоидными (имеют в своих клетках по 16 хромосом). Развитие яйцеклетки без оплодотворения носит название партеногенеза (от греческого parthenos — девственница), и, следовательно, трутни имеют партено-генетическое развитие. Генетиками было выяснено, что все наследственные различия между видами определяются не столько числом хромосом или их величиной и формой, сколько содержимым, которое в них находится. Это содержимое заключено в наследственных единицах — генах, контролирующих передачу потомству отдельных признаков и свойств. Гены располагаются в линейной последовательности вдоль всей хромосомы. Каждый ген занимает свое определенное место в одной хромосоме. Место его расположения в хромосоме определено термином локус (от латинского locus — место). Гены ответственны за все, что является у организмов врожденным и наследственным. Каждый из генов контролирует наследование одного, а иногда и нескольких признаков. Подобно хромосомам, они расположены попарно; с хромосомами они расщепляются в разные гаметы и затем вновь сходятся в процессе оплодотворения. Гены, расположенные в одних и тех же локусах гомологичных хромосом, — это гены-партнеры и они определяют развитие какого-либо признака у пчел. Гены-партнеры, дающие различный результат (см. Пример наследования окраски тела у пчел), называются аллеломорфами, или аллелями. Слово «аллеломорф» означает разно-форменный. Если число таких аллелей больше двух, то они образуют серию множественных аллелей. Каждая хромосома содержит только один из аллелей и потому любая диплоидная особь (матка, пчела) содержит два аллеля, в то время как гаплоидный трутень может иметь только один аллель какого-либо гена. Когда матка имеет два одинаковых аллеля в обеих гомологичных хромосомах, образующих гомологичную пару, то ее называют гомозиготной по данной паре аллелей. В случае же различающихся аллелей (имеют различное проявление признака) такая матка будет гетерозиготной по этой же паре аллельных генов. У трутня в связи с партеногенетическим развитием нет отца, в связи с чем у него только один набор хромосом и потому он не может быть гомозиготным или гетерозиготным. Особей, которые не имеют парных (гомологичных) хромосом, принято называть гемизиготными. Сумма всех генов организма в клетке, его наследственная конституция, получила название генотипа, а весь комплекс внешних признаков и свойств организма — морфологических, анатомических, физиологических, биохимических, инстинктивных и других, определяемый взаимодействием генотипа и внешней среды, называется фенотипом. Весь комплекс воздействий внешней среды на данный генотип носит название паратипа. Взаимоотношения генотипа и внешней среды можно выразить следующей формулой: генотип + паратип = фенотип. Каждая пчелиная семья имеет свой генотип и потому даже дочери от одних и тех родителей не бывают совершенно сходными между собой, всегда имеются определенные различия как между потомками, так и между родителями и их потомством. Еще большее разнообразие (изменчивость) можно заметить в стаде, или популяции. Если мы внимательно понаблюдаем на пасеке за пчелиными семьями, то очень скоро заметим, насколько они различаются друг от друга по интенсивности и характеру летной деятельности, по отношению их к окружающей среде. Такое явление носит название фенотипической изменчивости организмов. Фенотипическое разнообразие того или иного признака пчелиных семей характеризуется посредством определения границ изменчивости (лимитов), а также коэффициента вариации (Cv) (Коэффициент вариации есть отношение стандартного отклонения (а) к средней величине (М), выраженное в процентах. Коэффициент вариации позволяет сравнивать изменчивость различных признаков между собой независимо от единиц измерений ). Чем больше границы изменчивости, т. е. колебания от меньшего значения до большего (лимиты), и чем больше коэффициент вариации, тем больше изменчивость данного признака. В. Г. Кашковский (1977), изучая размеры трутневых ячеек на 40 семьях среднерусской породы, которым давал пустые рамки для отстраивания трутневых сотов, установил, что каждая пчелиная семья отстраивает разные по величине трутневые ячейки и эта разница довольно значительная. Например, пчелиная семья № 55 отстраивала трутневые ячейки со средней шириной 7,15 мм (колебания от 7,0 до 7,23), а семья № 13 — 6,31 мм (колебания от 6,11 до 6,40), т. е. разница между ними составила 13,3%. Многими исследователями было показано, что коэффициент вариации яйценоскости маток и силы пчелиных семей составляет 15—40%, а продуктивности по меду и воску — 20—50%. Наименьшую изменчивость имеют экстерьерные признаки у пчел (Cv = 1—6%) и масса маток, рабочих особей и трутней (Cv = 6—10%). Исследованиями Орловской опытной станции по пчеловодству было показано, что некоторые пчелиные семьи в условиях Орловской области на слабом медосборе собирали до 80 кг меда и отстраивали более 20 гнездовых рамок искусственной вощины, а яйценоскость отдельных маток была на уровне 3 тысяч яиц в сутки и выше (табл. 1). 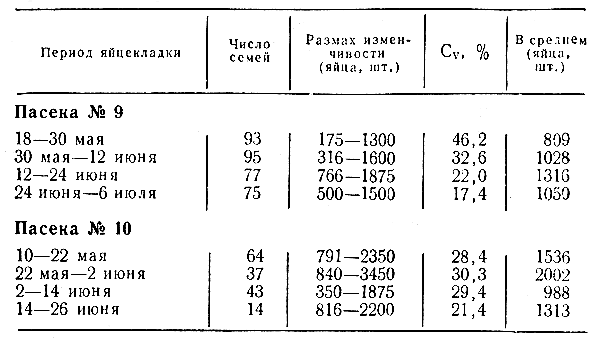 Таблица 1. Среднесуточная яйценоскость среднерусских маток Орловской популяции (по Н. Н. Гранкину) Согласно этим данным, наблюдаем изменчивость семей по яйценоскости маток как за определенный промежуток времени от минимума до максимума внутри всей группы, так и по периодам сезона. Если изменчивость по яйценоскости маток в первом случае можно квалифицировать как индивидуальную изменчивость (внутри группы), то во втором случае (по времени сезона) — как сезонную. Селекционеру пчел всегда приходится считаться как с индивидуальной изменчивостью пчелиных семей, которая дает ему материал для отбора лучших семей и их размножения, так и с сезонной, когда ему необходимо определенно решить, в какие сроки проводить наиболее объективный отбор лучших пчелиных семей. В целом можно сказать, что фенотипическое разнообразие по ведущим селекционным признакам у пчел достаточно высокое, что говорит о больших возможностях отбора по этим признакам. При выведении или улучшении определенных пород или линий животных селекционеру постоянно приходится иметь дело как с изменчивостью животных, откуда он берет необходимый материал (например, лучшие семьи по медопродуктивности), так и с наследственностью, строя таким образом селекционный процесс, чтобы успешно сохранить и закрепить найденное качество. Общая теория селекции основана на принципах, открытых и изученных генетикой, и состоит из нескольких разделов. Отбор по качественным (альтернативным) признакам. Первый из них элементарный менделизм, или по образному выражению А. С. Серебровского, — «арифметика селекции». Примером качественных признаков у пчел может служить окраска тела рабочих особей. При скрещивании тульских черных маток с итальянскими желтыми трутнями в потомстве получается преобладающее количество желтоокрашенных пчел и маток и черных трутней. Это означает, что желтая окраска тела у пчел доминирует (от латинского dominans — господствующий, главенствующий) над черным цветом тела, который по отношению к желтому будет рецессивным (от латинского recessus — отступление). Ген, проявление которого преобладает над проявлением другого, называется доминантным, а более слабый — рецессивным. Родители с черной и желтой окраской тела называются родительским поколением и сокращенно их принято обозначать символом — Р, а их детей (первое поколение) — символом F1, внуков (второе поколение)— F2 и т. д. Окраска тела (желтая и черная) обозначается как контрастные генетические признаки, обусловленные парой аллельных генов.
Символы ♂ (самец) и ♀ (самка), используемые биологами для обозначения половой принадлежности, заимствованы у астрономов, которые давно уже приняли копье в качестве символа планеты Марс, а зеркальце — в качестве символа планеты Венера. Поскольку у пчел, кроме яйцекладущих самок (маток), имеются еще и недоразвитые самки (рабочие особи), то их принято обозначать символом Сокращенными символами обозначают как проявившиеся внешние признаки, так и непроявившиеся признаки, обусловленные рецессивными генами, находящимися в генотипе животного. Так, прописной буквой Ж обозначают желтую окраску, строчной ж — черную. Из сказанного следует, что чистопородные среднерусские матки, пчелы и трутни не могут нести доминантного гена Ж (иначе они были бы желтой окраски) и потому они являются константными по рецессивному гену. Их потомство будет только черным: генетическая формула, или генотип их по окраске тела, будет — жж — у самок и ж — у трутней. Точно так же чистопородная итальянская матка и трутень будут иметь соответственно ЖЖ и Ж. Разумеется, что генетическая основа многих качественных признаков несколько сложнее, но в качестве простейшего приведенный пример, позволяющий уяснить основные генетические термины и принцип действия альтернативных признаков, вполне приемлем. Отбор по количественным признакам. Теория селекции, основанная на количественных признаках, допускает значительно большее применение математики; это уже «алгебра теории селекции». Для селекции сельскохозяйственных животных гораздо большее значение имеют признаки, которые определяются не одной парой генов, как в рассмотренном выше случае с качественными признаками, определяемыми парой аллельных генов, а большим числом однозначного действия неаллельных генов (иногда их больше сотни), которые ответственны за отдельный признак. Такие гены (полигены) называются множественными, или полимерными. Хорошей иллюстрацией количественного признака у медоносных пчел может служить их медопродуктивность, где при определенном среднем медосборе будут находиться как лучшие семьи, собравшие меда в 2 раза больше, чем средние семьи, так и худшие пчелиные семьи с очень низкой медопродуктивностью. Изменчивость в этом случае по медопродуктивности пчелиных семей становится непрерывной (в отличие от изменчивости качественных признаков) и различия между семьями, которые близки по своим результатам, можно установить только при помощи точных измерений. В связи с этим распределение пчелиных семей по медопродуктивности на графике будет представлять собой плавную кривую. Эта кривая получила в математике (математической статистике) наименование нормальной кривой. Кривая нормального распределения (рис. 1) отражает широко распространенные свойства вещей, подчиняясь замечательным закономерностям: чем ближе медопродуктивность пчелиной семьи к средней величине на данной пасеке (средняя медопродуктивность), тем чаще она встречается и, наоборот, по мере возрастания уклонения от средней (в обе стороны «плюс» и «минус») частота встречаемости падает в начале медленно и, наконец, спускается до нуля. Для селекционера всегда представляют большой интерес пчелиные семьи, уклоняющиеся в «плюс» и «минус» стороны от средней величины. Например, отбирается лучшая пчелиная семья с «плюс» медопродуктивностью или с «минус» ройливостью или злобливостью. 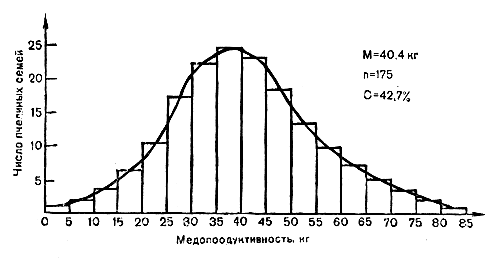 Рис. 1. Фенотипическое распределение группы пчелиных семей по медопродуктивности (средняя медопродуктивность 40,4 кг, интервал между классами 5 кг) В. В. Тряско (1958), изучая вопрос наследования длины хоботка рабочих особей при скрещивании пород пчел, контрастных по длине хоботка (серая горная кавказская и башкирская среднерусская породы пчел), выявила, что длина хоботка полностью наследуется по типу полигенного признака. Помеси первого поколения проявили промежуточное наследование, а возвратное скрещивание метисов на одну из пород приближало длину хоботков почти к исходным породам. Поскольку все важнейшие хозяйственно-полезные признаки медоносных пчел (медопродуктивность, зимостойкость, плодовитость и т. д.) имеют полигенное наследование, то отбор по количественным признакам является основной селекционной работой. Для этой категории признаков характерно то, что пчелиные семьи, имеющие одинаковый генотип, могут иметь различный фенотип и, наоборот, пчелиные семьи, имеющие одинаковые фенотипы (например, собравшие одно и то же количество меда), могут иметь различные генотипы. Допустим, что в весенний период на данной пасеке пчелиные семьи находятся в условиях, благоприятствующих их развитию (поддерживающий взяток, хорошее утепление, достаточная сила семей и т. д.). После того как семьи развились и достигли большой силы, пчеловод вывез основную часть семей сначала на липовый, а затем на гречишный медосбор и отобрал от них в конце сезона по 40 кг товарного меда. Остальные семьи он оставил на стационаре, рассчитывая с них взять такое же количество меда в среднем на семью на месте, где продолжался поддерживающий взяток. Эти семьи почти целиком изроилисъ и в результате не обеспечили себя кормовыми запасами на зиму, несмотря на то, что по наследственным качествам и возрасту маток были равнозначны основной части семей. В этом случае примерно одинаковые генотипы пчелиных семей имели различное фенотипическое проявление (различная степень ройливости и медопродуктивности) только из-за различий в условиях внешней среды, Если в первом случае (на кочевке) возможен отбор лучших семей, поскольку их генотипы полностью проявились в соответствующих условиях медосбора, то среди кочевавших семей отбор невозможен, хотя генотипы их равнозначны кочевавшим семьям. Определенными приемами (подсиливание, обогрев и т. д.) можно «придать» семье с плохим генотипом хорошее фенотипическое проявление, не уступающее лучшим пчелиным семьям с хорошими генотипами. Таким образом, при наследовании количественных признаков, когда одному фенотипу может соответствовать множество генотипов и, наоборот, сильно затрудняется суждение о наследственных задатках особи по ее фенотипу. Закон регрессии. Доказательством того положения, что фенотипические различия животных далеко не полностью совпадают с генотипическими, служат хорошо известные животноводам и селекционерам факты регрессии (возврата) потомства от отобранных лучших по фенотипу животных к средней величине для данной породы или данного стада. Первым, кто выявил явление частичного возврата потомства к средней величине, был Ф. Гальтон, который подошел к наследованию признаков статистически (Математический подход к изучению наследственности позволил Гальтону вместе с Пирсоном основать новую науку — биометрию (биологическую статистику) ), использовав большой материал, характеризовавший количественные признаки целых популяций, и вычислив связь между количественным выражением этих признаков у родителей и потомков. Разрабатывая материал о росте родителей и детей у английского населения, он установил закон регрессии, согласно которому отклонения родителей от среднего типа (в «плюс» и «минус» стороны) наследуются также их потомками, но не в полной мере: только 2/3 части уклонений от средней по росту наследуется потомками, а 1/3 часть уклонений имеет возврат к средней, или регрессию. Затем закон регрессии был проверен практически на всех сельскохозяйственных животных, в том числе и медоносных пчелах, и во всех случаях потомки от лучших родителей имели частичный возврат к средней величине.
|
 |
 |

ПОИСК:
|
 |
 .
.
© PASEKA.SU, 2001–2019
Хаустова Наталья разработка оформления
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paseka.su/ 'Paseka.su: Всё о пчеловодстве'
