 |
Новости |  |
|
Библиотека | |
|
Новые книги | |
|
Ссылки | |
|
Карта сайта | |
|
О нас | |
 |

Нервная системаКлеточная структура нервной ситемы. Нервная система состоит из тонких, не видимых простым глазом нервных клеток (в среднем менее 0,1 мм в диаметре). В нервной клетке различают расширенную часть с ядром — тело клетки — и несколько отростков, один из которых обычно значительно длиннее остальных; его называют нервным волокном (рис. 46, Л, нв). Отростки заканчиваются несколькими тоненькими веточками - нервными окончаниями. Нервные клетки своими окончаниями примыкают друг к другу, образуя единую систему. 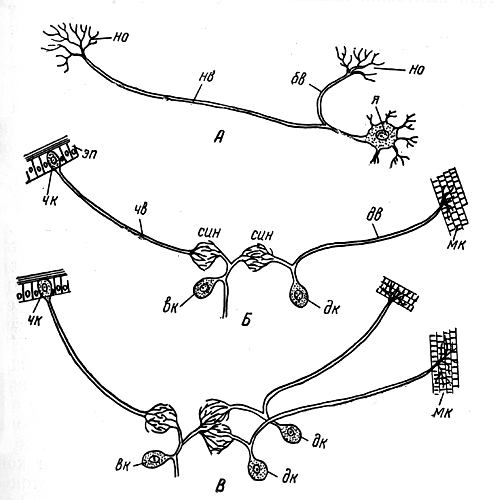 Рис. 46. Три типа нервных клеток и их связи (схема): А — двигательная нервная клетка; я — ядро; нв — волокно (длинный отросток, или аксон, нервной клетки); бв — боковая ветвь (коллатераль) нервного волокна; но — нервные окончания; Б — простая рефлекторная дуга, образованная чувствительной (чк), вставочной (вк) и двигательной (дк) нервными клетками; эп - эпидермис; чв — чувствительное волокно (волокно чувствительной нервной клетки); син - синапс; дв — двигательное волокно (волокно двигательной (чкл клетки); мк — мышечные клетки; В — чувствительная нервная клетка; (чк) - соединена вставочной нервной клеткой (вк) с боковыми ветвями двух мигательных нервных клеток (дк); мк — мышечные клетки Общая анатомия нервной системы. У низших многоклеточных животных (кишечнополостные) нервные клетки располагаются близко к поверхности тела. По мере прогрессивного развития животных некоторые из этих клеток погружались вглубь, сосредоточиваясь вдоль линии тела. В результате у всех высших животных нервная система подразделилась на периферическую и лежащую в глубине тела центральную. Клетки периферической нервной системы, называемые чувствительными нервными клетками, входят в состав органов чувств. В центральной нервной системе пчелы тела нервных клеток сконцентрированы в ганглиях (рис. 47, гн), как правило, по два ганглия на каждый сегмент соответственно левой и правой сторонам. Оба ганглия в сегменте соединены перемычкой из нервной ткани, называемой комиссурой. Продольные тяжи, связывающие между собой ганглии каждой стороны тела, получили название коннективов (рис. 47, кн). Ввиду того, что оба ганглия в сегменте сближены до соприкосновения, поперечная комиссура внешне становится невидимой, хотя коннективы остаются разделенными. Комиссуры и коннективы полностью свободны от тел нервных клеток и представляют собой пучки параллельных нервных волокон, клеточные тела которых находятся в ганглии. Нервные волокна начинают ветвиться лишь после вхождения в ганглий; там они вступают в контакт с окончаниями волокон других нервных клеток. В голове пчелы находится главная часть центральной нервной системы — мозг, отличающийся сложным внутренним строением. 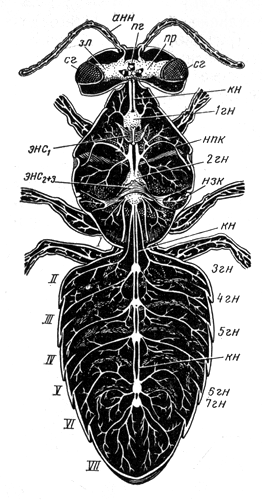 Рис. 47. Общий вид нервной системы пчелы-работницы: анн — антеннальный нерв; пр — протоцеребрум; зл — зрительная лопасть мозга; пг — простой глаз; сг — сложный глаз; гн — ганглий брюшной нервной цепочки; нпк — нерв переднего крыла; нзк — нерв заднего крыла; энс — эндостернит переднегруди; энс2+3 — эндостернит среднегруди и заднегруди; I — VII — брюшные сегменты; кн — коннективы С органами чувств и мышцами центральная нервная система соединена нервами, отходящими от ганглиев. Подобно комиссурам и коннективам, нервы слагаются из нервных волокон, покрытых общей оболочкой. Те нервные волокна, которые связывают центральную нервную систему с мышцами (рис. 46, Б, мк) и служат проводами для сигналов к их сокращению, называются двигательными. По волокну и всю клетку, которой оно принадлежит, называют двигательной, или мотонейроном. Тело мотонейрона находится в ганглии. Нервные волокна, подводящие к центральной нервной системе сигналы от органов чувств, получили название чувствительных. Чувствительное волокно отходит от тела чувствительной клетки, расположенного вдали от центральной нервной системы (около эпидермиса в органе чувств). Заканчивается чувствительное волокно в ганглии центральной нервной системы. Нервы бывают трех родов: чувствительные, двигательные и смешанные. В последнем случае нерв объединяет в себе как чувствительные, так и двигательные волокна. Поэтому в смешанном нерве сигналы передаются в двух противоположных направлениях: по чувствительным волокнам — от органов чувств к центру, по двигательным — от центра к мышцам; при этом, однако, и в чувствительной и в двигательной клетке сигнал распространяется от тела клетки к концевым разветвлениям волокна. В ганглиях между мотонейронами и окончаниями чувствительных волокон вклиниваются так называемые вставочные нервные клетки (рис. 46, 5, вк), не имеющие длинных отростков и не выходящие за пределы центральной нервной системы. Клеточные тела и нервные окончания располагаются в ганглии в определенном порядке. По периферии ганглия сосредоточены тела вставочных и двигательных клеток, а сердцевина заполнена сплетением концевых разветвлений отростков чувствительных, вставочных и двигательных клеток. Внутри ганглиев нет циркуляции гемолимфы: снабжение питательными веществами, равно как и удаление продуктов обмена, происходит через оболочку ганглия. Центральная нервная система. В центральной нервной системе пчелы различают головной мозг, состоящий из надглоточного и подглоточного ганглиев, и брюшную нервную цепочку (туловищный мозг) с двумя ганглиями в груди и пятью в брюшке (цв. табл. 1, рис. 1). Из всех ганглиев только надглоточный располагается дорсально по отношению к пищеварительному тракту; остальные находятся под кишечником на вентральной стороне тела. Надглоточный ганглий представляет собой важнейшую часть головного мозга. Он включает в себя три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум (прото-, дейто-, трито- от греческих слов «первый», «второй», «третий»; церебрум — по-латински «мозг»). Каждый отдел имеет симметричное строение, т. е. левая и правая его половины одинаковы. Протоцеребрум (рис. 48, пц) занимает верхнюю часть мозга. С боков от протоцеребрума отходят крупные зрительные лопасти, связанные со сложными (фасеточными) глазами. Снизу к нему примыкает дейтоцеребрум в виде двух грушевидных антеннальных лопастей, охватывающих с боков глотку; от каждой из них отходит крупный чувствительный нерв к органам чувств антенн и более мелкие двигательные нервы к мышцам антенн. За дейто-церебрумом лежит тритоцеребрум, но он настолько мал, что определить его положение можно только по месту отхождения лобных коннективов и верхнегубных нервов, которые выходят из-под антеннальных лопастей. Лобные коннективы связывают тритоцеребрум с лобным ганглием, расположенным кпереди от мозга; лобный ганглий посылает вперед, к мышцам, расширяющим цибарий, рединный лобный нерв, а назад — возвратный нерв, проходящий по верхней стенке кишечника. По очертаниям и размерам головной мозг пчелы-работницы, трутня и матки неодинаков. Мозг трутня в целом гораздо крупнее мозга матки и пчелы-работницы. Однако его массивность обусловлена главным образом большими размерами головы трутня и усиленным развитием зрительных лопастей. Последние по своей структуре и функции соответствуют не мозгу, а сетчатке глаза позвоночного животного. Что же касается собственно мозга, то у трутня он наименьший; фактически наибольшей величины он достигает у пчелы-работницы. Подглоточный ганглий (рис. 48, пгн) лежит в нижней части головы под глоткой и соединен с мозгом окологлоточными коннективами. У взрослой пчелы эти коннективы, однако, настолько коротки, что подглоточный ганглий кажется прикрепленным непосредственно к тритоцеребральной части мозга. Подглоточный ганглий — широкая, уплощенная масса нервной ткани, из которой выходят три пары крупных смешанных нервов к ротовым придаткам: мандибулярные, максиллярные и нижнегубные нервы. Кзади подглоточный ганглий отдает пару длинных коннективов, идущих к первому грудному ганглию (рис. 47). 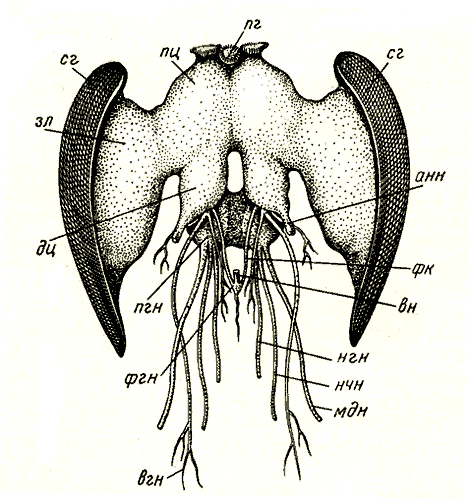 Рис. 48. Головной мозг пчелы-работницы с главными нервами головы (вид спереди): пг — простой глаз; сг — сложный глаз; зл — зрительная лопасть мозга; пц - протоцеребрум; дц — дейтоцеребрум (антеннальная лопасть мозга); анн — антеннальный нерв; пгн — подглоточный ганглий; фгн — фронтальный ганглий; фк — фронтальный коннектив; вн — возвратный нерв; нгн — нижнегубной нерв; нчн — нижнечелюстной нерв; мдн — мандибулярный нерв; вгн — верхнегубной нерв То обстоятельство, что подглоточный ганглий иннервирует три пары придатков, свидетельствует о том, что он представляет собой ганглии трех сегментов, объединившихся в одну массу. Брюшная нервная цепочка взрослой пчелы состоит из семи парных ганглиев (рис. 47, 1 гн — 1 гн) и их коннективов. Первый ганглий находится в переднегруди над стернитом этого сегмента; его основные нервы идут к первой паре ног. Второй ганглий в виде очень крупной нервной массы лежит впереди основания средних ног. Нервы от второго ганглия подходят к основанию обеих пар крыльев, к средним и задним ногам и к первому сегменту брюшка за стебельком. Второй ганглий — сложный, образовавшийся в результате объединения ганглиев четырех иннервируемых им сегментов; у личинки эти ганглии обособлены и расположены посегментно (рис. 32, Б, бнц). Следующие два ганглия пчелы находятся во втором и третьем сегментах брюшка, но, поскольку нервы каждого из названных ганглиев идут в следующий сегмент, эти ганглии принадлежат соответственно третьему и четвертому сегментам. Пятый ганглий находится в пятом брюшном сегменте и иннервирует его. Шестой ганглий расположен в шестом сегменте, но снабжает нервами и седьмой. Поэтому шестой ганглий сложный; у личинки он представлен двумя отдельными ганглиями (рис. 32, Б, бнц). Последний ганглий взрослой пчелы, лежащий в передней части седьмого сегмента, соответствует последнему ганглию личинки, который уже у нее состоит из слившихся ганглиев восьмого, девятого и десятого сегментов (рис. 32). Превращение простой структуры брюшной цепочки личинки в то состояние, которое свойственно взрослой пчеле, завершается на стадии куколки. Нервный импульс. Прежде считали, что сигналы, передающиеся по нервным волокнам, имеют электрическую природу. Дальнейшие исследования показали, что сигнал, или, как его обычно называют, нервный импульс, движется по нервному волокну гораздо медленнее, чем ток по проводам, а именно со скоростью от 7,5 до 90 м в секунду. Хотя это довольно большая скорость, все же она значительно уступает скорости движения электрического тока (160 000 км в 1 с). Нервный импульс представляет собой сложный комплекс химических и электрических явлений, разыгрывающихся в клеточной мембране нервного волокна. Клеточная мембрана — очень тонкий наружный живой слой цитоплазмы нервной клетки. Сверху она покрыта более плотной оболочкой клетки, представляющей собой неживое образование — продукт жизнедеятельности клетки. Внешняя поверхность мембраны нервного волокна заряжена положительно, а внутренняя — отрицательно. Напряжение между обеими поверхностями зависит от того, в какой мере мембрана проницаема для ионов, находящихся на ее внутренней и наружной поверхностях. Если она легкопроницаема, то ионы будут проходить сквозь нее и нейтрализовать друг друга, в силу чего напряжение уменьшится. Под влиянием внешних воздействий на органы чувств проницаемость мембраны чувствительного волокна в месте отхождения его от клеточного тела увеличивается, положительные и отрицательные ионы начинают взаимодействовать между собой, и напряжение в этом участке мембраны временно падает. Уменьшение напряжения в одном участке способствует увеличению проницаемости мембраны в соседнем участке нервного волокна. Таким путем вдоль всего нервного волокна распространяется волна снижения напряжения. Рефлекторная дуга и рефлекс. Импульсы следуют по чувствительному волокну один за другим с разной скоростью, в зависимости от силы внешнего воздействия на орган чувств. Дойдя до разветвления нервного волокна, импульсы одновременно попадают в несколько нервных окончаний. Клеточные оболочки, покрывающие как нервные волокна, так и клеточные тела, изолируют их друг от друга на всем протяжении, за исключением мест контакта концевых окончаний, получивших название синапсов (от греческого слова «смыкать») (рис. 46, Б, сип). Дойдя до конца волокна, импульс в большинстве случаев не в состоянии перейти на концевые разветвления примыкающей вставочной клетки и стимулирует здесь выделение органического вещества ацетилхолина. Под влиянием последнего во вставочной нервной клетке заново возникает импульс, быстро распространяющийся по ее отростку. В двигательную клетку импульсы передаются через синапс между окончаниями вставочной клетки и окончаниями боковой ветви двигательного волокна (рис. 46). Нервные окончания двигательного волокна, разветвляющегося в мышце, под влиянием нервных импульсов выделяют химическое вещество, вызывающее сокращение мышечных волокон. У насекомых каждый мотонейрон контролирует почти все мышечные волокна, а каждое мышечное волокно связано со многими мотонейронами, из которых одни генерируют слабые токи и вызывают медленные сокращения мышечного волокна, другие же служат источниками гораздо более сильных токов и возбуждают больше волокон. Кроме того, имеются тормозные нервные клетки, которые могут подавлять действие других мотонейронов. Благодаря такой организации обеспечивается тонкая регуляция движений при малом общем количестве мотонейронов и мышечных волокон. У позвоночных на каждое мышечное волокно приходится только одно нервное окончание и возможен лишь один вид возбуждения мышц, зато сами мышцы делятся на быстрые и медленные. Путь нервных импульсов от чувствительной клетки через вставочную к двигательной называют рефлекторной дугой, а ответ, в данном случае сокращение мышцы, — простым рефлексом. Прохождение импульсов по рефлекторной дуге в общих чертах легко можно представить, но в действительности в центральной нервной системе концевые разветвления множества чувствительных волокон, коротких отростков вставочных и двигательных клеток запутанно переплетаются друг с другом. Благодаря вставочным клеткам число проводящих путей, по которым распространяются импульсы, почти не ограничено. Автономность ганглиев брюшной нервной цепочки. Каждый сегмент тела обладает значительной степенью автономии. Вероятно, каждый сегмент имеет свой собственный дыхательный центр, контролирующий движение расположенных в нем дыхалец, и в состоянии произвести такие движения, в которые не вовлекаются другие сегменты. Изолированный последний сегмент брюшка осуществляет рефлекс внедрения и изъятия жала. Грудной отдел пчелы, с которым связано большинство ее движений, будучи изолирован, совершает ряд действий, свойственных пчеле, а именно: хождение, схватывание, взмахивание крыльями. Координация движений, связанных счисткой антенн, может осуществляться и после разрушения надглоточного ганглия. Подглоточный ганглий, по-видимому, равноценен ганглиям туловищного мозга в том отношении, что он контролирует движение ротовых придатков. Свойственные каждому сегменту (или группе их) определенные стереотипные (т. е. проявляющиеся в неизменном виде) движения обеспечиваются постоянными рефлекторными путями на уровне соответствующего ганглия. Величина ответа определяется поступающими в этот ганглий импульсами с чувствительных клеток органов чувств. Однако сигналы от органов чувств поступают одновременно в высшие центры и на другие уровни центральной нервной системы. Высшие отделы контролируют автономные рефлекторные системы посредством возбуждающих и тормозных волокон, передавая приказы двух типов: стимулирующие их деятельность или прекращающие ее. Главный координирующий центр головного мозга — стебельчатые тела, а наиболее дифференцированный и сложно устроенный его отдел — протоцеребрум. Большую часть протоцеребрума между зрительными лопастями и занимают стебельчатые тела — парное образование, называемое иначе грибовидными телами (рис.49, А, ст). Каждое тело имеет на верхнем конце два расширения в форме чаши, поддерживаемой толстой ножкой, которая на нижнем конце раздваивается на доли, или «корешки». Одна доля заворачивает вперед в лобную область мозга, другая идет вниз и к середине, почти встречаясь с соответствующей долей противоположной стороны (рис. 49, Б). Каждая чаша образована нервными волокнами и как бы наполнена массой мелких вставочных клеток. Отростки клеток идут в ножку, но прежде отдают ветвь в чашу; несколько групп волокон проходит от одной чаши к другой. К стебельчатым телам подходят отростки вставочных клеток из зрительных лопастей, из дейтоцеребрума, из подглоточного ганглия. Они вступают в стебельчатые тела у основания чаши и через переднюю корешковую долю. Из стебельчатых тел нервные отростки выходят в составе нижней корешковой доли и направляются к так называемому центральному телу протоцеребрума, к двигательным центрам (скоплениям мотонейронов) антеннальных мышц, к двигательным центрам тритоцеребрума и подглоточного ганглия. 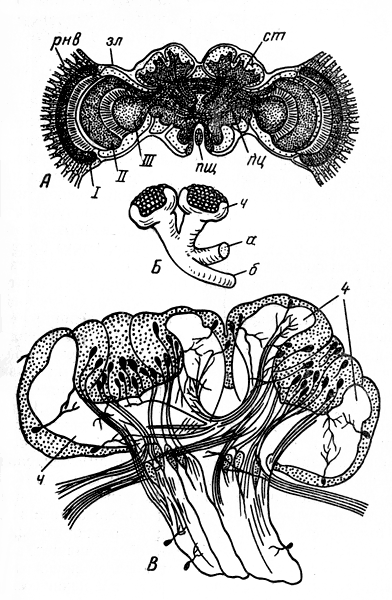 Рис. 49. Внутреннее строение головного мозга (схема): А - фронтальный (параллельный лобной поверхности) срез через мозг пчелы-работницы; рнв - ретинальный нерв; зл - зрительная лопасть мозга; I - ганглиозная пластинка зрительной лопасти; II - наружная медулляоная пластинка зрительной лопасти; III - внутренняя медуллярная пластинка зрительной лопасти; ст - стебельчатое (грибовидное) тело; пц - протоцеребрум; пщ - пищевод; Б - стебельчатое тело правой стороны (вид спереди); ч - чаша стебельчатого тела; а - передний корешок стебельчатого тела; б - срединный корешок стебельчатого тела; В - срез через стебельчатое тело протоцеребрума пчелы-работницы (ч - чаша) Помимо своей роли главных координирующих центров мозга, стебельчатые тела обеспечивают возможность наиболее сложных форм поведения пчелы. Искусственным воздействием на нервные клетки слабым электрическим током можно имитировать ту реакцию, которая в естественных условиях возникает под влиянием нервных импульсов: в исследуемую область вводят электроды из вольфрамовой, стальной или платиновой проволоки диаметром 10—30 мкм (10—30 тысячных долей миллиметра). Электроды изолированы на всем своем протяжении за исключением самого кончика. Электрическим раздражением отдельных участков стебельчатых тел у пчелы-работницы можно вызвать разнообразные формы поведения: отрыгивание пищи из медового зобика, чистку, агрессивное поведение, строительную деятельность. Если одна форма поведения вызвана, то любая другая, до тех пор наблюдавшаяся, подавляется. Очевидно, это обусловлено возникновением конфликта в мотонейронах, участвующих в осуществлении обеих реакций. Конфликт разрешается путем подавления одной из них. Деятельность мозга не исчерпывается процессами возбуждения и торможения, создания условных рефлексов и их разрушения. Эти процессы являются базой для работы специальной системы управления целенаправленными действиями — психики. Именно эта система обеспечивает координацию мышечной активности. Психика не синоним понятия «сознание». Любой высокоразвитый организм выказывает свою психику. По поведению млекопитающего, птицы, рептилии, рыбы, насекомого можно видеть, что данное существо куда-то стремится, чего-то боится, в состоянии как-то ориентироваться в обстановке и даже «прогнозирует» возможные ситуации и свои действия в них. На психическом уровне, т. е. на уровне высшего управления организмом, поведение строится на основе учета информации о ведущей потребности организма в данный момент. Одновременно оценивается ситуация в среде в отношении того, насколько она способствует удовлетворению потребностей. При этом постоянно привлекается прошлый опыт, зафиксированный в клетках мозга. Все это происходит до размышления, когда речь идет о человеке, и без размышления, если имеют в виду животных, которые не умеют размышлять. Молодая пчела-работница с функционирующими гипофарингеальными железами, обнаружив в гнезде в восковой ячейке сота здоровую личинку младшего возраста, снабжает ее молочком. Та пчела-работница, у которой хорошо развиты восковые железы, испытывает неудовлетворенность и беспокойно блуждает по гнезду до тех пор, пока не натолкнется на строительную гирлянду других особей и не присоединится к ним.
|
 |
 |

ПОИСК:
|
 |
© PASEKA.SU, 2001–2019
Хаустова Наталья разработка оформления
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paseka.su/ 'Paseka.su: Всё о пчеловодстве'
