 |
Новости |  |
|
Библиотека | |
|
Новые книги | |
|
Ссылки | |
|
Карта сайта | |
|
О нас | |
 |

Формы взаимосвязи в пчелиной семьеДля согласования в пчелином сообществе разделения труда нет надобности в специальном координирующем центре. Формы активности каждого индивида предопределены его наследственностью. Загрязненная ячейка для пчелы-работницы служит стимулом к ее вычищению. Нуждающаяся в пище личинка — стимул к ее кормлению для пчелы с активными гипофарингеальными железами, т. е. в возрасте не менее трех дней; при этом личинкам разных стаз питание поставляется неодинаковое. Вполне развитая личинка — стимул для запечатывания ячейки. Необходимый для строительной деятельности воск лучше всего секретируется восковыми железами пчелы-работницы с восьмого по восемнадцатый день ее жизни. Таким образом, пчелы-работницы удовлетворяют те потребности пчелиной семьи, которые служат безусловными стимулами их врожденного поведения. Время проявления разных форм поведения определяется физиологическим состоянием пчел и окружающей их обстановкой. Каждая пчела-работница сама отыскивает соответствующие стимулы, т. е. сама информирует себя о потребностях сообщества, и действует в любой обстановке как его представитель, в меру своих возможностей. Если разделение труда в пчелиной семье является автоматическим результатом самостоятельной активности тысяч пчел-работниц, то для других целей они вступают в определенные взаимоотношения друг с другом. Общность запаха. Основная предпосылка существования пчелиной семьи состоит в том, что пчелы-работницы не покидают ее, возвращаясь после кратковременных вылетов в родное гнездо. Индивиды других семей нетерпимы в данной семье. «Удостоверением», которое должно быть предъявлено каждой пчелой, входящей в улей, служит специфический запах этой семьи, удерживающийся в опушении пчелы. Запах семьи сложный. Он слагается из запахов нектара и пыльцы, собранных пчелами семьи. Если в конце лета для обеспечения пчелиных семей дополнительными запасами корма на зиму подкармливать их сиропом, которому искусственно придан аромат, например, перечной мяты, то всем пчелиным семьям будет сообщен один запах, и сторожа не смогут отличить обитателей своих ульев от посторонних. В специфический запах пчелиной семьи, возможно, входит составной частью и запах матки, поскольку пчелу-работницу из безматочной семьи другая пчелиная семья принимает. Феромоны. Для существования пчелиной семьи крайне важно, что каждая пчела, заметив опасность, прежде чем вступить в борьбу, подает сигнал тревоги, который быстро передается от одной пчелы к другой. Наблюдать подобную систему сигнализации в действии лучше всего к вечеру или в холодный день, когда у летка остаются единичные сторожевые пчелы. Если надавить на спинку такой пчелы, она тут же поднимет кончик брюшка, обнажит жалоносный аппарат, и тогда между волосками мягкой перепонки, соединяющей продолговатые пластинки жала, начнет проступать жидкость, запах которой и служит сигналом тревоги (рис. 78). Иногда эта жидкость собирается в капельку на конце жала (запах, поднимающий тревогу, не идентичен запаху яда). Побегав в этой позе перед летком, пчела затем устремляется в улей и вскоре из него высыпает масса пчел-работниц, готовых к нападению. Нападению подвергаются темные подвижные предметы, особенно с шершавой или ворсистой поверхностью. Такая удивительно быстрая отзывчивость пчелиного сообщества на сигнал об опасности, полученный всего-навсего от одного его индивида, достойна восхищения. Здесь проявляется существенная черта общественного образа жизни: для проявления некоторых врожденных форм поведения вовсе не обязательно каждому индивиду самому сталкиваться с безусловным раздражителем этой поведенческой реакции — достаточно сигнала о наличии его от другого индивида.  Рис. 78. В ответ на механическое раздражение (сдавливание спинки) пчела подает ароматический сигнал об опасности Насонова железа и железа в основании жалоносного аппарата, запах секрета которой воспринимается пчелами как сигнал тревоги, представляют собой экзокринные железы. В отличие от эндокринных желез (внутренней секреции), продукты которых (гормоны), выделяемые во внутреннюю среду организма, осуществляют регуляцию и координацию функций других органов и тканей, экзокринные железы выделяют свои продукты в проток, ведущий на поверхность тела. Вещества, вырабатываемые экзокринными железами, получили название феромонов. Попадая во внешнюю среду, феромоны влияют на поведение других особей того же вида. Известно, что информация может передаваться от одного животного к другому посредством зрительных и слуховых воздействий; феромоны же представляют собой средство передачи сообщений путем воздействия на органы обоняния или вкуса (хеморецепторы). Феромоны выделяются не только пчелами-работницами, но и маткой, в основном ее мандибулярными железами. Но если феромоны пчелы-работницы воздействуют через хеморецепторы на центральную нервную систему другой соплеменницы и вызывают немедленную поведенческую реакцию последней, то феромоны матки обладают более медленным действием и вызывают у пчел-работниц цепь физиологических явлений, приводящих к подавлению у них развития яичников. Благодаря тому, что матка свободно передвигается по сотам в области ячеек с расплодом, она обходит практически всех находящихся там молодых пчел-работниц. Возможность периодически ощущать своими антеннами маточные феромоны создает пчелам-работницам необходимый фон для их нормальной деятельности. Пищевые контакты. В пчелином сообществе широко распространены пищевые контакты между его индивидами. В ответ на вытягивание хоботка одной пчелой другая отрыгивает каплю пищи. Обе врожденные формы поведения — «просить» корм у пчелы-работницы своей семьи и в свою очередь отрыгивать его, когда «просят» другие, — свойственны пчелам-работницам с первого дня жизни. На основании следующего опыта можно судить, какое место занимает эта форма взаимосвязи в жизни пчелиной семьи. Шести пчелам-сборщицам дали возможность собрать 20 мл раствора сахара, содержащего радиоактивный фосфор. Через 4 ч 62% сборщиц и 16-21% всех пчел-работниц данной семьи, насчитывавшей 25500 особей, были радиоактивны. По истечении 48 ч были радиоактивны все личинки. Первые передачи пчелой-разведчицей ульевым пчелам найденного ею в поле и собранного нектара информируют пчелиную семью о наличии в окрестностях улья корма, его качестве и о том, на цветках с каким ароматом его следует искать. Влияние информации последнего рода на поведение лётных пчел по отношению к растениям разных видов особенно эффектно показывает опыт, проведенный в местности, где в естественном травяном покрове много цикламен, а в усадьбах широко распространены флоксы. На кормовом столике пчелам-сборщицам предлагают небольшой букетик цикламен. Предварительно на каждый цветок наносят по капле сиропа. Вскоре дикорастущие цикламены, как и неподслащенные букеты из них, начинают привлекать множество пчел-работниц. В то же самое время к флоксам пчелы-работницы не проявляют ни малейшего интереса. Если же теперь заменить цикламены на кормовом столике букетиком флоксов, наполнив перед тем трубочки его венчиков сиропом, то уже спустя несколько минут картина меняется. Пчелы, обычно никогда не посещающие флоксов вследствие недоступности для них нектара этого растения, находящегося на дне слишком длинной трубки венчика его цветка теперь летят только на флоксы, как бы вовсе не замечая цикламен. Аромат нектара. Аромат посещенных пчелой-сборщицей цветков источает как принесенный и раздаваемый ею в гнезде нектар, так и она сама. Опушение пчелы из многочисленных тонких хитиновых волосков прекрасно приспособлено для впитывания цветочных ароматов. Последние удерживаются на пчеле лучше, чем на стекле, металле, вате; даже внешний покров других насекомых уступает в этом отношении покрову пчелы-работницы. Если искусственно придать сиропу и опушению пчелы разный запах, то можно выяснить, что важнее в качестве средства информации: аромат ли нектара или же благоухание самой пчелы. Если, например, предложить пчелам-работницам невдалеке от их улья на кормовом столике сироп, настоенный на цветках флокса, но при этом поставить пчел в такие условия, что набрать сироп с ароматом флокса они смогли бы не иначе, как только взобравшись на цветки цикламена (рис. 79), то в таком случае пчелы-работницы, вылетевшие по сигналу подопытных сборщиц, будут посещать и флоксы и цикламены. Если же, однако, увеличить расстояние от улья до кормового столика, например до 600 м, то главное значение приобретает запах нектара, в данном случае флоксов. Новички устремляются теперь преимущественно на флоксы. Запах последних не может улетучиться при транспортировке нектара в медовом зобике, тогда как аромат, передавшийся от цветков цикламена опушению пчелы, за время ее дальнего полета в какой-то степени выветривается. 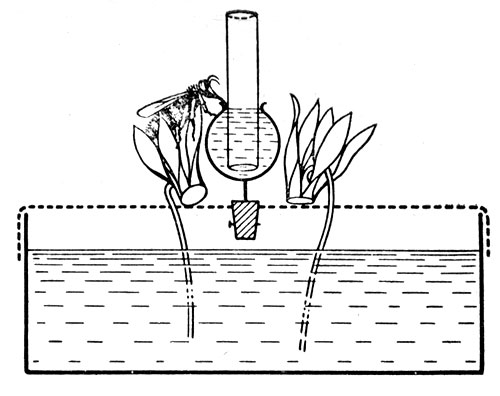 Рис. 79. Пчела набирает настоянный на цветках флокса сахарный сироп, взобравшись на цветки цикламена (по Фришу) В отличие от сборщиц нектара у пчел-работниц, приносящих в гнездо цветочную пыльцу в виде двух пыльцевых комочков, укрепленных в корзиночке голени на каждой задней ноге, именно душистая обножка служит средством информации о виде посещенных ими растений. Обладательница обножек не спешит выложить их из корзиночек в подходящую для хранения белкового корма ячейку, но задерживается среди своих соплеменниц, и последним предоставляется возможность воспринять и хорошенько запомнить своеобразие аромата принесенной пыльцы. Аромат пыльцы и лепестков цветка заметно различается. Следующие опыты доказывают, что сигнальное значение имеет аромат пыльцы, а не лепестков, хотя последний, несомненно, передался опушению сборщицы пыльцы, когда она находилась в цветках. Одну группу пчел-работниц приучают добывать пыльцу в цветках колокольчика, а другую — в цветках шиповника, причем их букеты помещают на разные кормовые столики, изрядно удаленные друг от друга. По прошествии некоторого времени оба букета убирают, и лет пчел прекращается. После перерыва на тот же столик, где прежде были обычные колокольчики, выставляют такие цветки колокольчика, из которых удалены тычинки и пестик, а на их место вложены тычинки шиповника (рис. 80). Первая пчела, прилетевшая к столику, найдя на привычном месте цветки колокольчика, собирает подмененную пыльцу. Однако по возвращении в улей ее окружают соплеменницы не из ее группы, а те, что прежде собирали пыльцу шиповника. Появление в улье запаха шиповника служит для них сигналом к вылету. И они летят к столику, где собирали пыльцу шиповника и местоположение которого освоили в предыдущих полетах. Но, увы... их столик пуст. Подобный же результат получается и при замене тычинок в цветках шиповника пестиками из цветков колокольчика с налипшей на их столбики пыльцой. 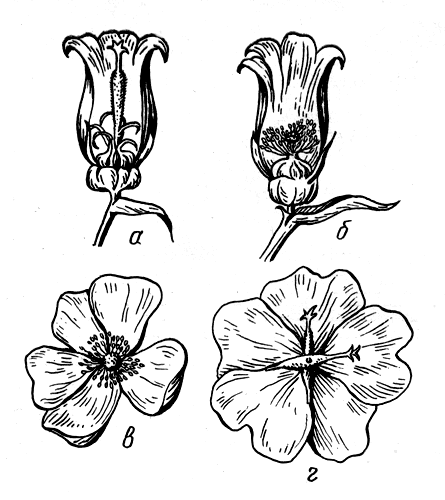 Рис. 80. Опыт с замещением тычинок в цветках одного вида растения тычинками другого вида: а — цветок колокольчика; б — цветок колокольчика с тычинками шиповника; в — цветок шиповника; г — цветок шиповника с двумя пестиками колокольчика, обсыпанный пыльцой последнего (по Фришу) Танец пчел-сборщиц. Иногда пчела-сборщица, еще не успев войти в улей, передает доставленный ею нектар пчелам-работницам, встретившим ее на прилетной доске. Одновременно принимающие нектар 4—5 пчел стоят перед ней веером, устремив вытянутые хоботки в отрыгиваемую ею из медового зобика каплю. Через 20—30 с передача корма внезапно прекращается резким поспешным поворотом сборщицы брюшком к пчелам — приемщицам нектара. Быстрыми, мелкими шажками устремляется она по прилетной доске в направлении, откуда прилетела с нектаром, нашедшим такой повышенный спрос в ее семье, но вдруг, как бы спохватившись, возвращается назад, затем опять повторяет своеобразный бег в том же направлении и снова возвращается, как будто ее что-то удерживает, и она не решается улетать. Между тем приемщицы нектара не спешат уходить в гнездо — сборщица продолжает оставаться в центре их внимания. Но теперь их интересуют ее движения, и, раздавшись вширь, они располагаются вокруг нее. Одни из них выглядят пассивными зрителями, другие пытаются следовать за пчелой-сборщицей. Внимательно приглядевшись к последней, можно заметить, что во время пробегов по прямой линии она вся дрожит, причем всего сильнее колеблется из стороны в сторону самый кончик ее брюшка. К нему и протягивают приемщицы нектара свои антенны. Сделав несколько таких пробегов с виляниями в правильном ритме и под одним углом к солнцу, сборщица нектара теперь сама обращается за кормом к одной из приемщиц и, получив его и почистив хоботок и глаза, взмывает ввысь. Особенно красивое зрелище предстает перед глазами наблюдателя, когда вышеописанные ритмичные движения, получившие название танца, совершает сборщица пыльцы с полными корзиночками цветной обножки. Частота виляний во время пробегов в танцах постоянна и составляет 13 полных циклов в 1 с. Закончив первый пробег, танцующая пчела тут же возвращается точно на то место «танцевальной площадки», где она его начинала, и, заняв исходную позицию, повторяет пробег в прежнем направлении. Фазу танца, во время которой пчела возвращается в исходную позицию перед очередным пробегом, называют поворотом. Поскольку пробеги в танце невелики и редко превышают длину тела самой пчелы-работницы более чем вдвое, танцовщице для возвращения в начальный пункт нет надобности преодолевать значительное расстояние по окружности, как это часто изображают авторы популярных очерков. Поворачиваясь, пчела движется наподобие стрелки часов. Но в отличие от часовой стрелки ось вращения танцовщицы, проходящая вблизи ее головного конца, не фиксирована, а слегка перемещается, описывая кривую малого радиуса. Иными словами, танцовщица осуществляет поворот в результате сложного движения, состоящего из двух простых: вращательного и поступательного. Точки поверхности головы, спинки и брюшка танцовщицы описывают при ее повороте разные траектории. На рисунке 81 в качестве геометрического образа танцовщицы взята ее продольная ось. Чем короче пробег, тем слабее проявляется в следующем за ним повороте поступательное движение и тем меньше площади требуется для танца. Танец с хорошо выраженным пробегом получил название восьмерочного (рис. 81, А). Ему противопоставляют круговой танец, в котором преобладает вращательное движение, пробег же ничтожно мал, а площадь танца минимальна (рис. 81, Б). Повороты по часовой стрелке чередуются в танце с поворотами в противоположном направлении. Один полный цикл танца складывается из трех элементов: поворота влево, пробега и поворота вправо. 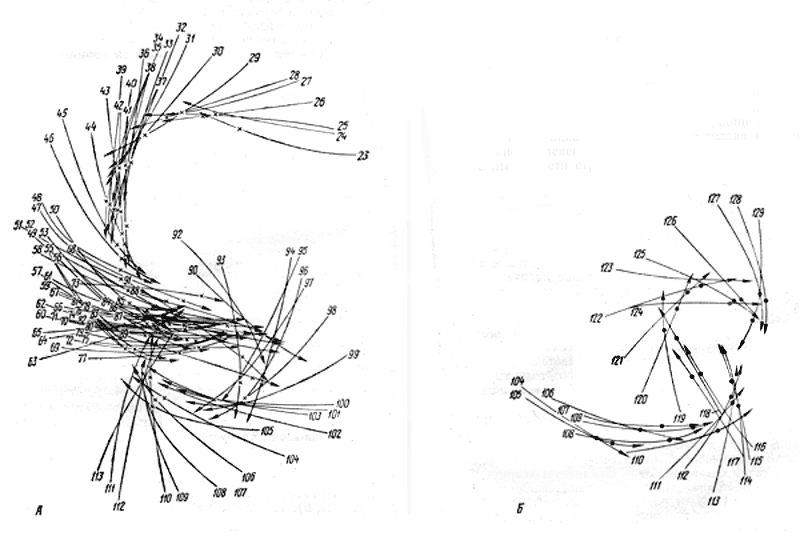 Рис. 81. Танец пчелы на соте в гнезде (фрагмент). Последовательные положения продольной оси тела танцующей пчелы через равные промежутки времени: А — восьмерочный танец: поворот влево (23—46), прямолинейный пробег с виляниями (47-89), поворот вправо (90-113); источник корма в 700 м от гнезда; Б — круговой танец: поворот влево (104—115), прямолинейный пробег (116—118), поворот вправо (119—129); источник корма в 5 м от гнезда. Обратите внимание, что площадь кругового танца значительно меньше площади восьмерочного танца. Чертеж воспроизводит танцы, заснятые на киноленту (увеличено; чертеж автора) Обычно танец совершается пчелами-сборщицами в гнезде на сотах и, стало быть, в темноте. Если изолировать наблюдательный улей от попадания в него солнечных лучей (установив его в темном помещении и сделав вывод леткового отверстия наружу с помощью коридорчика), то при красном свете можно убедиться, что и на сотах направление каждого последующего пробега сборщицы в танце совпадает с первым. Когда пчела-сборщица движется под постоянным углом к солнцу на прилетной доске, она ориентируется по солнцу или по направлению поляризации рассеянного света. На вертикальной же поверхности сотов впотьмах ориентировка по зрению исключена. Выдержать одно направление в этих условиях пчеле-сборщице позволяют органы гравитационного чувства. Ее голова и брюшко, подвижно сочлененные с грудным отделом, под действием силы тяжести стремятся занять положение равновесия, подобно тому как маятник при отклонении его внешней силой около точки подвеса стремится вернуться в вертикальное положение. В зависимости от положения пчелы на вертикальном соте разные участки поверхности сочленения в области шеи и брюшного стебелька будут испытывать разное давление. Последнее воспринимается находящимися здесь чувствительными зонами органов гравитационного чувства (см. рис. 58). Каждому положению пчелы на вертикальной или наклонной поверхности соответствует свое особое распределение давления среди многочисленных осязательных волосков, образующих чувствительные зоны. Если находящийся в темном помещении наблюдательный улей при красном свете повернуть так, чтобы сот расположился в горизонтальной плоскости, органы гравитационного чувства перестанут служить пчеле измерительным прибором. В каком бы направлении ни двигалась пчела по горизонтальной поверхности, распределение давления в области шеи и брюшного стебелька останется неизменным. Хорошо ориентируясь в темноте на вертикальных сотах, пчела утрачивает эту способность на горизонтальной поверхности, так как изменение направления движения больше не отражается на сигнализации от органов чувств. На горизонтальной поверхности пчеле для ориентировки необходим свет. А без света пчела-сборщица хотя и продолжает повторять пробеги с виляниями, но специфическая особенность всей совокупности последовательных пробегов — постоянство их направления — утрачивается: теперь что ни пробег, то новое направление. Но стоит лучам света упасть на горизонтальный сот, как восстанавливается правильный танец. Упорядоченный, с единым направлением всех пробегов танец легко получить на горизонтальном соте, если стоящий под открытым небом наблюдательный улей, заселенный небольшой пчелиной семьей, положить плашмя (над сотом оставляют одну стеклянную дверцу). Если кормовой столик, посещаемый танцующей пчелой, находится к западу от улья, а солнце светит с юга, то лететь к столику по прямому курсу пчеле удается в силу того, что на протяжении всего полета она стремится удержаться в таком положении, чтобы солнце сильнее всего освещало тот омматидий левого глаза, который составляет прямой угол с продольной осью тела пчелы. Благодаря этому линия полета пчелы образует с направлением от улья к солнцу прямой угол (90°); иными словами, пчела располагается продольной осью своего тела правее солнца на 90°. Точно так же встанет пчела на освещенном горизонтальном соте по отношению к солнцу перед началом пробега (рис. 82). Ее пробег с виляниями укажет поэтому на запад. Когда же улью будет возвращено его нормальное, вертикальное положение, и притом так, чтобы к югу, т. е. к солнцу, сот был обращен той стороной, где пчела-сборщица передает корм и танцует, значение солнца для нее как единственного ориентира во время танца полностью сохраняется. Во время пробега она займет положение на 90° правее солнца, т. е. снова укажет на запад (во время пробега, направленного в восточную сторону, пчела тоже двигалась бы под прямым углом к солнцу, однако не правее его, а левее). Примечательно, что на вертикальном соте пчела освещается солнцем не сбоку, как в полете, а со спинной стороны. Вследствие этого максимальное освещение получают иные омматидий. Боковой омматидий левого глаза, служивший пчеле при полете компасной стрелкой, указывавшей на солнце, на вертикальном соте как бы передает свою функцию омматидию на верхней стороне того же глаза.  Рис. 82. Направление прямолинейного пробега в танце на горизонтальной поверхности (справа) совпадает с направлением предшествовавшего полета танцовщицы к источнику корма (по Фришу) У каждого омматидия есть свой напарник. Совершая пробег, танцовщица становится на соте так, чтобы наибольшее количество света попадало именно в соответствующий омматидий-напарник. При полете к постоянной цели пчела-сборщица руководствуется движущимся ориентиром — солнцем, характер перемещения которого она освоила на опыте и благодаря чувству времени и памяти может запомнить его положение на небосводе в любое время суток. Если со времени предыдущего вылета пчелы за кормом, например, в западном направлении, прошло полчаса, то, выйдя из улья для нового полета, она встает по отношению к солнцу не так, как прежде, а под углом меньше первого на столько угловых градусов, на сколько сместилось солнце к западу за эти полчаса (в среднем на 7,5°, для умеренной зоны). Вследствие изменения угла линии полета по отношению к солнцу в точном соответствии с угловой скоростью перемещения солнца направление полета пчелы в отношении стран света остается одним и тем же. Точно так же остается неизменным и направление прямолинейного пробега на освещенном горизонтальном соте. Но когда пчела возвращается с кормом в темный улей с вертикальными сотами, она не в состоянии более ориентировать свои движения по солнцу. Выбрать направление прямолинейного пробега в танце она здесь может благодаря гравитационному чувству. И хотя направление полета за кормом определялось без участия органов гравитации, направление прямолинейных пробегов в темноте на вертикальной поверхности обнаруживает явную зависимость от положения источника корма. Две системы органов чувств — зрительная и гравитационная, контролирующие направление движения пчелы, одна на свету, а другая в темноте на вертикальной поверхности, — оказываются взаимосвязанными. Каждому углу падения солнечных лучей на сложный глаз пчелы во время ее полета за кормом соответствует свое, особое направление прямолинейного пробега в гравитационном поле. За полетом пчелы-работницы в направлении к солнцу автоматически следует пробег на соте прямо вверх. Подобно тому как на свету перед взлетом пчела для определения полета постоянно как бы измеряет угол от направления к солнцу, принимая его за начало отсчета, в гравитационном поле в темноте нулевой линией ей служит направление вертикально вверх, т. е. диаметрально противоположное направлению силы земного притяжения. При кормовом столике, расположенном к западу от улья, и солнечном освещении с южной стороны, когда линия полета пчелы ориентирована под углом 90° правее солнца, пчела, танцуя впотьмах на вертикальном соте, направит свой прямолинейный пробег под углом 90° правее вертикали. Направление прямолинейного пробега на протяжении дня испытывает закономерные изменения. По мере того как солнце клонится к западу, прямолинейный пробег перестает совпадать с горизонталью, а все больше приподнимается над ней, становясь все более отвесным, т. е. угол между вертикалью и прямолинейным пробегом делается более острым. Когда же кормовой столик и солнце окажутся на одной линии к западу от улья, пчела во время виляния побежит по соту точно вверх (рис. 83). В замене на соте направления к солнцу постоянным направлением вверх отражается истинное положение вещей: не Солнце премещается, а Земля вращается вокруг Солнца, и вместе с Землей меняется относительно Солнца направление от улья к источнику корма. 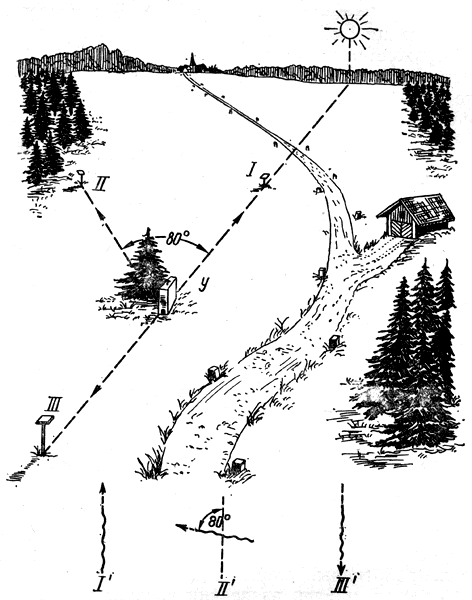 Рис. 83. Три разных направления прямолинейного пробега в танцах пчел на вертикальном соте впотьмах: У — улей; I, II, III — кормовые столики в трех различных направлениях; I', II', III' — соответствующие направления прямолинейного пробега в танце (по Фришу) Если, открыв дверцу наблюдательного улья, дать возможность солнечным лучам осветить танцующую на вертикальном соте в темноте пчелу, последняя мгновенно изменит направление своего пробега. В этих условиях гравитационное чувство утрачивает всякое значение для ориентировки танца. В отличие от прямых солнечных лучей, рассеянного света недостаточно, чтобы свести на нет роль гравитационного чувства. Сигнализация от омматидиев глаз, воспринимающих направление поляризации света неба, не подавляет сигнализацию от органов гравитационного чувства, характер которой сложился еще при полете пчелы к кормовому столику. В условиях освещения рассеянным светом пчела одновременно стремится ориентировать прямолинейный пробег и по направлению поляризации света неба и в соответствии с сигнализацией от органов гравитации. В результате пробег происходит в промежуточном направлении, занимающем точно среднее положение между направлением пробега в темноте и направлением пробега на вертикальном соте, освещенном солнцем (Фриш, 1962). Пчелы-работницы, заинтересованные в сборе того корма, который найден разведчицей, принимают информацию, содержащуюся в ее танце, и разыскивают источник корма под тем углом к солнцу, который соответствует направлению в ее танце прямолинейного пробега с виляниями. 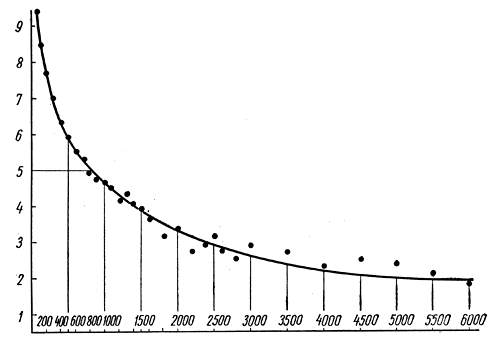 Рис. 84. Зависимость частоты прямолинейных пробегов в танце пчелы-сборщицы от расстояния до источника корма. Абсцисса — расстояние до источника корма в метрах. Ордината — число прямолинейных пробегов за 15 секунд. График построен на основе 3885 танцев пчел. Каждая точка графика — средняя многих наблюдений. Пчелы, посещавшие кормовой столик, отстоявший от улья на 100 м, успевают за четверть минуты совершить от 9 до 10 прямолинейных пробегов и столько же полных циклов танца (поворот вправо + пробег + поворот влево). При расстоянии до кормового столика 200 м число пробегов за то же время снижается до 7; при 1 км — составляет 41/3, а при 6 км — только 2. Не все точки совпадают с кривой. Одна из причин отклонения — влияние на скорость танца направления ветра. Встречный ветер при полете к кормовому столику оказывает то же действие, что и удлинение пути, — он замедляет темп танца; попутный же ветер, напротив, убыстряет танец. Оценка пчелой удаленности источника корма основана на ее способности измерять усилия, затраченные ею на то, чтобы добраться до него (по Фришу)  Кроме направления полета, танец содержит информацию о расстоянии до источника корма. Если при постепенном удалении кормового столика наблюдать за танцем посещающих его пчел и подсчитывать, сколько прямолинейных пробегов успевают они сделать за один и тот же отрезок времени, то окажется, что с увеличением расстояния частота пробегов снижается. Так, для пчел-сборщиц крайнской популяции получены следующие значения (округленные) числа прямолинейных пробегов за четверть минуты танца при разной удаленноcти от улья кормового столика: Темп танца замедляется неравномерно, и поэтому график зависимости частоты пробегов от расстояния проходит не по прямой линии (рис. 84). При одном и том же расстоянии темп танца пчел-работниц разных рас неодинаков (табл. 4). 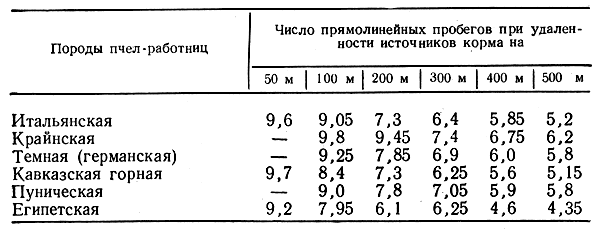 Таблица 4. Скорость танца (число прямолинейных пробегов за четверть минуты) пчел-работниц шести пород Снижение числа пробегов в танце в единицу времени по мере увеличения дальности полета танцовщицы объясняется удлинением самого пробега, а вместе с ним и всего цикла танца (рис. 85). Напротив, чем ближе цель полета, тем короче прямолинейный пробег. А при дальности полета 50 м пробег так мал, что становится трудным различить его простым глазом. В этом случае в движениях танцовщицы преобладают повороты, и танец становится круговым. 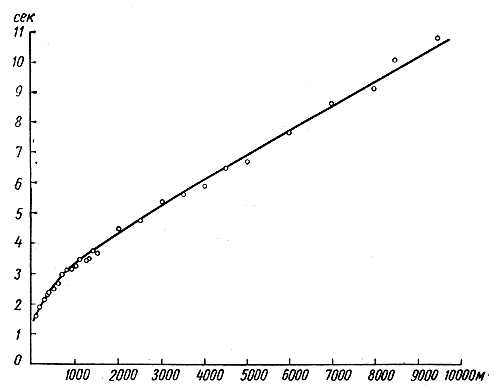 Рис. 85. Зависимость между продолжительностью одного цикла танца и расстоянием до источника корма. Абсцисса — расстояние до источника корма в метрах, ордината — продолжительность одного полного цикла (поворот вправо + прямолинейный пробег с виляниями + поворот влево) танца в секундах. График построен на основе наблюдений за 6267 танцами (по Фришу) Следовательно, направление прямолинейного пробега в танце отражает направление полета, а в длине пробега зашифровано его расстояние до источника корма. Танцовщица информирует об этом тех пчел-работниц, которые заинтересовались образцом найденного ею в природе корма и готовы лететь на его сбор. Помимо маршрута полета, танцовщица сообщает танцем о степени продуктивности и качестве источника корма. Танец может длиться от нескольких секунд до 1—3 мин. Чем изобильнее источник корма, тем продолжительнее танец. Предполагают, что сигналом высокого качества корма служит прерывистый звук, издаваемый танцовщицей во время прямолинейного пробега. Его можно услышать с помощью медицинского стетоскопа, приблизив последний расширенным концом к танцовщице. Воспроизводится звук мышцами крыла. По своей частоте, составляющей 250 колебаний в секунду, он не отличается от звука летящей пчелы. Не воспринимая звуковых колебаний воздуха и оставаясь индифферентными даже к громкому шуму, пчелы-работницы обладают высокой чувствительностью к звукам, распространяющимся по твердому субстрату. Резонансные колебания сотов, возникающие под действием звука, издаваемого танцовщицей, передаются ногам окружающих ее пчел-работниц и почти наверняка воспринимаются их субгенальными органами. Принося сироп средней концентрации, пчела-сборщица может тоже танцевать, но в этом случае она совершает прямолинейный пробег с виляниями беззвучно. Следующий опыт показывает, насколько точно воспринимают молодые пчелы-работницы темп танца или продолжительность прямого пробега с виляниями, сигнализирующими об удаленности источника корма. На различных расстояниях по прямой линии от улья до источника корма, посещавшегося мечеными пчелами, и несколько дальше размещали пахучие подставочки без корма. Если пчелы, отыскивая на показанном танцовщицей расстоянии запах, пролетали неподалеку от пахучих подставочек, то последние привлекали их. Пчелы подлетали к подставочкам, кружились над ними и в конце концов опускались на них; там их и подсчитывал наблюдатель. На рисунке 86 изображены графически результаты двух таких опытов. В первом испытании источник корма находился на расстоянии 750 м, во втором — 2000 м от улья. Числа на графике показывают, как много пчел, отыскивающих сироп, посетило во время наблюдения подставочку на данном расстоянии. Из графика видно, что сигнализация танцовщицей расстояния вполне точна и хорошо воспринимается другими пчелами-работницами. 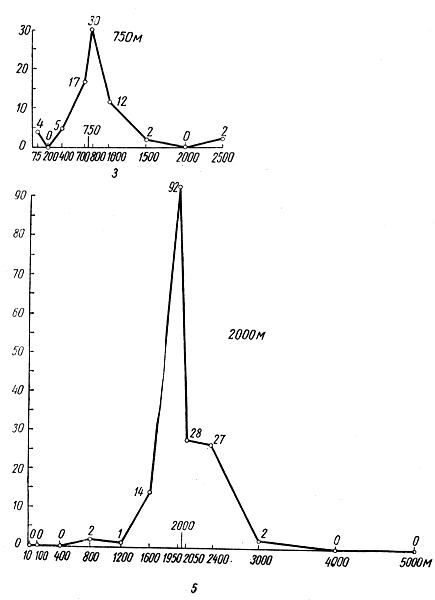 Рис. 86. Результаты двух экспериментов по изучению способности пчел воспринимать дальность полета до источника корма из сообщения пчелы-танцовщицы: вверху — удаленность кормового столика от улья 750 м, продолжительность наблюдений - 1 час 30 минут, внизу - удаленность кормового столика 2000 м, продолжительность наблюдений 3 часа. Абсцисса — расстояние между ульем и пахучими подставочками (м), ордината — число немеченных пчел зафиксированных на пахучих подставочках (по Фришу) Для выяснения, в какой степени молодые пчелы выдерживают направление, сообщенное им танцовщицей, ставили множество опытов; результаты одного из них графически представлены на рисунке 87. В 250 м от улья несколько меченых пчел получали корм на подставочках, снабженных лавандовым маслом. На расстоянии 200 м от улья веерообразно были расположены семь таких пахучих подставочек без корма: одна из них сходилась на прямой линии между ульем и местом, где пчелы собирали корм, три — влево от этой линии, а остальные три — вправо. Угол между линиями составлял 15°. Пчел, прилетевших на разные подставочки в течение часа, соответственно было 8, 13, 58, 132, 37, 7 и 3. Иными словами, подавляющее большинство пчел (58+1324-37 = 227, или 88%) не отклонялось от верного направления больше чем на 15°. Хотя пчелы-работницы и ориентировались по запаху, воспринятому ими от танцовщицы, но искали они этот запах в определенном направлении. 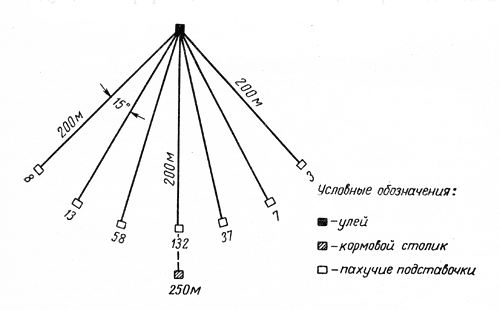 Рис. 87. «Веерный» эксперимент по изучению способности молодых пчел воспринимать направление к источнику корма, сообщенное им пчелой-танцовщицей; числа у подставочек обозначают количество прилетевших немеченых пчел (по Фришу) Разведчица танцует в кругу пчел-работниц, испытывающих потребность в ее информации о маршруте полета. Как собираются к танцовщице ее соплеменницы, объединяемые одним желанием? Если понаблюдать за поведением разведчицы в гнезде после ее первых прилетов с ношей нектара, то можно убедиться, что танцевать в это время она не расположена. Она снует по гнезду и предлагает пчелам-работницам дегустировать открытый ею в природе источник медосбора. И лишь в том случае, если этот корм найдет спрос в пчелиной семье, то около разведчицы будут постепенно собираться молодые пчелы-работницы, готовые его добывать. Вот к этим жаждущим информации соплеменницам разведчица и обращает свой символический «рассказ» о маршруте полета к источнику корма, причем в танце содержится и точная характеристика его направления и количественный показатель необходимых энергетических затрат. Благодаря информации разведчиц соплеменницы пчелиного сообщества становятся осведомленными о тех явлениях внешнего мира, которые они непосредственно не воспринимали своими органами чувств. Следовательно, жизненный опыт одних индивидов пчелиного сообщества передается другим. И если первые обучались по способу проб и ошибок, то вторые освобождаются от затрат энергии на это обучение. Последнее оказывается для них излишним. Запомнив инструкции, полученные в гнезде от разведчицы, они напрямик летят за кормом. Проявляющаяся здесь особая форма обучения называется инструктивным обучением. Танец при роении. Сигнализицая с помощью танцев наблюдается в пчелином сообществе не только на сотах в гнезде, но и на роевой грозди. Здесь информационное содержание танца связано не с источником корма, а с местом, где надлежит пристать отделившемуся рою. Роение — единственная форма распространения медоносной пчелы. Оно способствует сохранению вида. Поэтому отыскание роем хорошего места для поселения имеет важное значение для переживания пчелиным сообществом неблагоприятного периода. Вскоре после того как роевые пчелы по выходе из материнского гнезда сформируют неподалеку от него роевую гроздь, на последней можно наблюдать танцующих пчел. Но теперь это не сборщицы корма, а разведчицы, отыскавшие подходящее место для поселения и сигнализирующие об этом временно привившемуся рою. Разведчицами становятся опытные пчелы-сборщицы старшего возраста. Поскольку разведчицы отыскивают жилище одновременно в разных местах, их танцы вначале сигнализируют о нескольких возможных жилищах. В танце каждой разведчицы отражается расстояние до найденного ею места и направление к нему. Прежде чем рой отправится к месту поселения, танцы большинства разведчиц становятся сходными, и рой летит к лучшему из всех найденных жилищ. Разведчицы, разыскавшие особенно благоприятное место, танцуют гораздо дольше и энергичнее по сравнению с теми, которые нашли менее подходящее место. Поэтому пчелы-работницы роя сильнее всего привлекаются к лучшему из найденных жилищ. Пчелы, повторявшие за танцовщицами их движения, покидают гроздь и летят на место, обозначенное танцем. По возвращении к рою они также начинают танцевать, вследствие чего воздействие на рой становится более интенсивным. Те разведчицы, которые нашли худшие места, изменяют свои побуждения, если рядом с ними более энергично танцуют пчелы, нашедшие лучшее место. Под влиянием неистовых танцовщиц первые сами летят к найденному другими разведчицами лучшему жилищу и только после ознакомления с ним меняют характер своего танца, тоже начиная сигнализировать о наилучшем месте. В то время как для пчел-сборщиц танец продолжительностью в 2 мин является редкостью, роевые пчелы могут танцевать беспрерывно целыми часами, причем угол между направлением прямого пробега и вертикалью изменяется в соответствии с солнечным углом. Пчелы, танцующие на вертикальной поверхности роевой грозди с теневой стороны, одновременно ориентируются и по поляризованному свету и по направлению силы тяжести. Пристальным наблюдением за танцами роевых пчел можно предсказать направление, в котором окончательно полетит рой, а также дальность его полета, достигающую иногда нескольких километров. Межвидовые различия танца. Танцы свойственны не только виду Apis mellifera L., но и остальным трем видам рода Apis. Танец индийской пчелы отличается более медленным темпом, но такая его особенность не выходит за пределы различий между породами внутри вида. Что же касается карликовой пчелы, то в отношении условий, необходимых для передачи информации о направлении и расстоянии, здесь обнаруживаются своеобразные черты более примитивного характера. Пчелы-сборщицы этого вида танцуют на верхней площадке сота. Сот у карликовой пчелы охватывает опору своей верхней, расширенной удлиненными ячейками для складывания запасов корма частью. Освещенные солнцем пчелы-сборщицы совершают на верхней горизонтальной поверхности его пробег с виляниями в направлении, совпадающем с направлением к источнику корма, напоминая танцы рабочих особей медоносной пчелы на прилетной доске. Если отрезать ветку с подвешенным к ней сотом и повернуть ее так, чтобы «танцевальная площадка» приняла вертикальное положение, все танцовщицы мгновенно прекращают танцевать. Они устремляются кверху и, отыскав малейшую горизонтальную площадку, снова возобновляют танцы. Если же горизонтальную площадку накрыть миниатюрной двускатной стеклянной крышей и таким образом лишить пчел возможности находить ее, то, как правило, они не танцуют. Немногие из них пытаются танцевать на вертикальной поверхности, однако в таком случае форма танцев утрачивает всякое соответствие с местоположением источника корма. Гигантская пчела занимает промежуточное положение между медоносной пчелой и карликовой по способу передачи информации. Пчелы-сборщицы этого вида совершают танцы на вертикальной поверхности. Сот у гигантской пчелы прикреплен к ветке верхним краем и лишен горизонтальной площадки. Перевод солнечного угла в угол к направлению силы тяжести у гигантской пчелы совершается по тому же принципу, что и у медоносной и индийской пчел. Но в отличие от последних гигантской пчеле для выполнения ориентированных танцев на вертикальном соте необходимо видеть небо, подобно карликовой пчеле на горизонтальной площадке. Более примитивный способ информации о найденном источнике корма у карликовой пчелы по сравнению с остальными видами рода Apis сам по себе все же представляет высокоразвитую форму взаимоотношений между индивидами сообщества. Симптом, сигнал, знаковая система. Пчелы-работницы создают в пчелиной семье условия, необходимые матке для выполнения ею своей функции. Они кормят ее, поддерживают в гнезде оптимальный микроклимат и строят ячейки, в которые она откладывает яйца. В свою очередь, пчелы-работницы нуждаются в феромонах матки, не являющихся для ни:: пищевыми веществами. Пчелы-работницы испытывают органическую потребность в воздействии маточных феромонов на свои хемо-рецепторы. От характера этого воздействия зависит физиологическое состояние их яичников и их поведение. Пчелы-работницы и матка образуют единую систему. Посредством феромонов матки в этой системе поддерживается подвижное равновесие. Феромоны матки служат здесь постоянным показателем ее состояния. В отличие от матки пчелы-работницы выделяют феромоны лишь в особых случаях, причем феромоны одних особей служат ориентиром или стимулом для целесообразной поведенческой реакции других. Феромоны пчел-работниц, следовательно, играют роль сигнала, тогда как феромоны матки, следует рассматривать как симптом. Найденный разведчицей и принесенный ею в улей на пробу нектар, скажем, цветков липы — тоже сигнал. Что же касается танцев пчел-работниц, то их следует отнести к средствам коммуникации более высокой категории по сравнению с сигналом. Танцы свидетельствуют о том, что пчелы-работницы в состоянии различать свои бесконечно разнообразные полеты по их общим элементам, направлению и протяженности. Выделить эти общие элементы невозможно без специальной знаковой системы. Такая система в виде языка танцев и выработалась в процессе эволюции медоносной пчелы.
|
 |
 |

ПОИСК:
|
 |
© PASEKA.SU, 2001–2019
Хаустова Наталья разработка оформления
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://paseka.su/ 'Paseka.su: Всё о пчеловодстве'
